Transpiració vegetal

- L’aigua es transporta passivament a les arrels i després al xilema.
- Les forces de cohesió i adhesió fan que les molècules d’aigua formin una columna al xilema.
- L’aigua es mou del xilema a les cèl·lules mesofil·les, s’evapora de les seves superfícies i surt de la planta per difusió a través dels estomes


La transpiració és el procés de moviment de l'aigua a través d'una planta i la seva evaporació a partir de parts aèries, com fulles, tiges i flors. L’aigua és necessària per a les plantes, però només s’utilitza una petita quantitat d’aigua captada per les arrels per al creixement i el metabolisme. Entre el 97-99,5% restant es perd per transpiració i gutació.[1] Les superfícies de les fulles estan farcides de porus anomenats estomes ("estoma" singular), i en la majoria de les plantes són més nombroses a la part inferior del fullatge. Els estomes estan vorejats per cèl·lules de protecció i les seves cèl·lules accessòries estomals (conegudes juntes com a complex estomàtic) que obren i tanquen el porus.[2] La transpiració es produeix a través de les obertures estomatals i es pot considerar com un "cost" necessari associat a l'obertura dels estomes per permetre la difusió del diòxid de carboni de l'aire per a la fotosíntesi. La transpiració també refreda les plantes, canvia la pressió osmòtica de les cèl·lules i permet el flux massiu de nutrients minerals i aigua de les arrels als brots. Hi ha dos factors principals que influeixen en la velocitat del flux d’aigua des del sòl fins a les arrels: la conductivitat hidràulica del sòl i la magnitud del gradient de pressió a través del sòl. Aquests dos factors influeixen en la velocitat del flux massiu d’aigua que es mou des de les arrels fins als porus estomàtics de les fulles a través del xilema.[3]
El flux massiu d’aigua líquida des de les arrels fins a les fulles es condueix en part per acció capil·lar, però principalment per diferències de potencial hídric. Si el potencial hídric de l’aire ambiental és inferior al potencial hídric de l'espai aeri foliar del porus estomàtic, el vapor d’aigua viatjarà pel gradient i es mourà de l'espai aeri foliar a l'atmosfera. Aquest moviment redueix el potencial hídric de l'espai aeri de les fulles i provoca l'evaporació de l’aigua líquida de les parets cel·lulars del mesòfil. Aquesta evaporació augmenta la tensió sobre els meniscos de l’aigua a les parets cel·lulars i disminueix el seu radi i, per tant, la tensió que s’exerceix sobre l’aigua de les cèl·lules. A causa de les propietats cohesives de l'aigua, la tensió viatja a través de les cèl·lules de la fulla fins al xilema de la fulla i de la tija, on es crea una pressió negativa momentània a mesura que l'aigua treu el xilema de les arrels.[4] A mesura que l'evaporació es produeix a la superfície de la fulla, les propietats d’adhesió i cohesió funcionen de forma conjunta per extreure molècules d’aigua de les arrels, a través del teixit del xilema i de la planta a través dels estomes.[5] En plantes i arbres més alts, la força de la gravetat només es pot superar amb la disminució de la pressió hidroestàtica (aigua) a les parts superiors de les plantes a causa de la difusió de l’aigua dels estomes a l’ atmosfera. L'osmosi absorbeix l'aigua a les arrels i els nutrients minerals dissolts viatgen amb ella a través del xilema.
La teoria de la tensió de cohesió explica com les fulles treuen aigua pel xilema. Les molècules d’aigua s’uneixen o presenten cohesió. Quan una molècula d’aigua s’evapora de la superfície de la fulla, tira de la molècula d’aigua adjacent, creant un flux continu d’aigua a través de la planta.[6]
Etimologia
[modifica]Podem veure la història de la paraula transpiració quan la desglossem en trans, un substantiu llatí que significa "a través" i espiració, que prové del verb llatí spīrāre, que significa "respirar". El sufix de ció afegeix el significat "l'acte de", de manera que podem veure que la transpiració és, literalment, "l'acte de respirar a través", que identifica clarament l'emissió de vapor de les fulles de les plantes. La transpiració és vital per a l'ecologia terrestre i s’està reduint per la nostra delmació dels boscos.
Acció capil·lar
[modifica]L'acció capil·lar és el procés d’un líquid que flueix en espais estrets sense l’ajut de les forces externes com la gravetat o fins i tot en oposició a aquestes. L'efecte es pot observar en la preparació de líquids entre els pèls d'un pinzell, en un tub prim, en materials porosos com el paper i el guix, en alguns materials no porosos com la sorra i la fibra de carboni liquada, o en una cèl·lula biològica. Es produeix a causa de les forces intermoleculars entre el líquid i les superfícies sòlides circumdants. Si el diàmetre del tub és prou petit, la combinació de la tensió superficial (que és causada per la cohesió dins del líquid) i les forces adhesives entre el líquid i la paret del contenidor actuen per propulsar el líquid.[7]
Regulació
[modifica]Les plantes regulen la velocitat de transpiració controlant la mida de les obertures estomals. La taxa de transpiració també està influenciada per la demanda evaporativa de l'atmosfera que envolta la fulla, com ara la conductància de la capa límit, la humitat, la temperatura, el vent i la llum solar incident. Juntament amb factors sobre el terreny, la temperatura i la humitat del sòl poden influir en l'obertura estomàtica[8] i, per tant, en la velocitat de transpiració. La quantitat d'aigua perduda per una planta també depèn de la seva mida i de la quantitat d'aigua absorbida a les arrels. La transpiració explica la major part de la pèrdua d’aigua d’una planta per les fulles i les tiges joves. També serveix per refredar evaporativament les plantes, ja que l’aigua que s’evapora emporta energia calorífica a causa de la seva gran calor latent de vaporització de 2260 kJ per litre.
| Funció | Efecte sobre la transpiració |
|---|---|
| Nombre de fulles | Més fulles (o espines o altres òrgans fotosintetitzadors) significa una superfície més gran i més estomes per a l’intercanvi gasós. Això provocarà una major pèrdua d’aigua. |
| Nombre d'estomes | Més estomes proporcionaran més porus per a la transpiració. |
| Mida de la fulla | Una fulla amb una superfície més gran transpirarà més ràpidament que una fulla amb una superfície més petita. |
| Presència de cutícula vegetal | Una cutícula cerosa és relativament impermeable a l’aigua i al vapor d’aigua i redueix l'evaporació de la superfície de la planta excepte a través dels estomes. Una cutícula reflectant reduirà l'escalfament solar i l’augment de temperatura de la fulla, ajudant a reduir la velocitat d’evaporació. Les petites estructures semblants als cabells anomenades tricomes a la superfície de les fulles també poden inhibir la pèrdua d’aigua creant un entorn d’alta humitat a la superfície de les fulles. Aquests són alguns exemples de les adaptacions de plantes per a la conservació de l'aigua que es poden trobar en molts xeròfits. |
| Subministrament de llum | La taxa de transpiració està controlada per l’obertura estomàtica i aquests petits porus s’obren especialment per a la fotosíntesi. Tot i que hi ha excepcions (com la fotosíntesi nocturna o CAM), en general, un subministrament de llum afavorirà els estomes oberts. |
| Temperatura | La temperatura afecta la velocitat de dues maneres: 1) Un augment de l'evaporació a causa d'un augment de la temperatura accelerarà la pèrdua d'aigua. 2) La disminució de la humitat relativa fora de la fulla augmentarà el gradient de potencial hídric. |
| Humitat relativa | L'entorn més sec proporciona un gradient de potencial d’aigua més fort i, per tant, augmenta la velocitat de transpiració. |
| Vent | En aire quiet, l'aigua perduda a causa de la transpiració es pot acumular en forma de vapor a prop de la superfície de la fulla. Això reduirà la taxa de pèrdua d’aigua, ja que el gradient de potencial hídric de l’interior a l'exterior de la fulla és lleugerament inferior. El vent bufa bona part d’aquest vapor d’aigua a prop de la superfície de la fulla, fent que el gradient de potencial sigui més pronunciat i acceleri la difusió de les molècules d’aigua a l’aire circumdant. Fins i tot en el vent, pot haver-hi una certa acumulació de vapor d’aigua en una fina capa límit d’aire en moviment més lent al costat de la superfície de la fulla. Com més fort sigui el vent, més fina tendrà a ser aquesta capa i més inclinat serà el gradient de potencial hídric. |
| Subministrament d'aigua | L'estrès hidric causat pel subministrament restringit d'aigua del sòl pot provocar un tancament estomàtic i reduir les taxes de transpiració. |
-
 L'efecte de la temperatura sobre la velocitat de transpiració de les plantes.
L'efecte de la temperatura sobre la velocitat de transpiració de les plantes. -
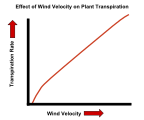 L'efecte de la velocitat del vent sobre la velocitat de transpiració de les plantes.
L'efecte de la velocitat del vent sobre la velocitat de transpiració de les plantes. -
 L'efecte de la humitat sobre la velocitat de transpiració de les plantes.
L'efecte de la humitat sobre la velocitat de transpiració de les plantes.




Durant una temporada de creixement, una fulla transpirarà moltes vegades més aigua que el seu propi pes. Un acre de blat de moro (4.047 m².) desprèn uns 3.000-4.000 galons (11.400-15.100 litres) d’aigua cada dia i un gran roure pot transpirar 40.000 galons (151.000 litres) a l’any. La relació de transpiració és la proporció de la massa d’aigua transpirada a la massa de matèria seca produïda; la proporció de transpiració dels conreus tendeix a caure entre 200 i 1000 (és a dir, les plantes de conreus transcendeixen entre 200 i 1000 kg d’aigua per cada kg de matèria seca produïda).[9]
Les taxes de transpiració de les plantes es poden mesurar mitjançant diverses tècniques, inclosos potòmetres, lisímetres, poròmetres, sistemes de fotosíntesi i sensors de flux de saba termomètrics. Les mesures dels isòtops indiquen que la transpiració és el component més gran de l’ evapotranspiració.[10] L'evidència recent d'un estudi global [11] d'isòtops estables d'aigua mostra que l'aigua transpirada és isotòpicament diferent de les aigües subterrànies i els rierols. Això suggereix que l'aigua del sòl no està tan ben barrejada com s'assumeix àmpliament.[12]
Les plantes del desert tenen estructures especialment adaptades, com ara cutícules gruixudes, zones de fulles reduïdes, estomes enfonsats i pèls per reduir la transpiració i conservar l’aigua. Molts cactus realitzen la fotosíntesi en tiges suculentes, en lloc de fulles, de manera que la superfície del brot és molt baixa. Moltes plantes del desert tenen un tipus especial de fotosíntesi, anomenada metabolisme àcid de les crassulàcies o fotosíntesi CAM, en què els estomes es tanquen durant el dia i s’obren a la nit quan la transpiració serà menor.
Cavitació
[modifica]Per mantenir el gradient de pressió necessari perquè una planta es mantingui sana, han de prendre contínuament aigua amb les seves arrels. Han de ser capaç de satisfer les demandes d’aigua perduda a causa de la transpiració. Si una planta és incapaç d’aportar prou aigua per romandre en equilibri amb la transpiració, es produeix un esdeveniment conegut com a cavitació.[13] La cavitació és quan la planta no pot subministrar el seu xilema amb aigua adequada, de manera que en lloc d’omplir-se d’aigua, el xilema comença a omplir-se de vapor d’aigua. Aquestes partícules de vapor d’aigua s’uneixen i formen bloquejos dins del xilema de la planta. Això evita que la planta pugui transportar aigua per tot el seu sistema vascular.[14] No hi ha cap patró aparent d’on es produeixi la cavitació al llarg del xilema de la planta. Si no es cuida eficaçment, la cavitació pot fer que una planta arribi al seu punt de marciment permanent i mor. Per tant, la planta ha de tenir un mètode per eliminar aquest bloqueig de cavitació o ha de crear una nova connexió de teixit vascular a tota la planta.[15] La planta ho fa tancant els estomes durant la nit, cosa que atura el flux de transpiració. Això permet que les arrels generin més de 0,05 mPa de pressió, la qual cosa fa possible la destrucció del bloqueig i reomplir el xilema amb aigua, reconnectant amb el sistema vascular. Si una planta no pot generar la pressió suficient per erradicar el bloqueig, ha d’evitar que el bloqueig s’estengui amb l’ús de peres de fossa i després crear un nou xilema que pugui tornar a connectar el sistema vascular de la planta.[16]
Els científics han començat a utilitzar la ressonància magnètica (RM) per controlar l'estat intern del xilema durant la transpiració, de manera no invasiva. Aquest mètode d’imatge permet als científics visualitzar el moviment de l’aigua a tota la planta. També és capaç de veure en quina fase es troba l’aigua mentre es troba al xilema, cosa que permet visualitzar esdeveniments de cavitació. Investigadors botànics van poder veure que al llarg de 20 hores de llum solar, més de 10 vasos de xilema van començar a omplir-se de partícules de gas cavitant-se. La tecnologia de ressonància magnètica també va permetre veure el procés mitjançant el qual es reparen aquestes estructures de xilema a la planta. Després de tres hores a la foscor, es va veure que el teixit vascular es subministrava amb aigua líquida. Això va ser possible perquè a la foscor els estomes de la planta estan tancats i la transpiració ja no es produeix. Quan s’atura la transpiració, les bombolles de cavitació són destruïdes per la pressió generada per les arrels. Aquestes observacions suggereixen que les ressonàncies magnètiques són capaces de controlar l'estat funcional del xilema i permet als científics veure els esdeveniments de cavitació per primera vegada.[15]
Referències
[modifica]- ↑ Sinha, Rajiv Kumar. Modern Plant Physiology (en anglès). CRC Press, 2004. ISBN 978-0-8493-1714-9.
- ↑ Freeman, Scott. Biological Science (en anglès). Prentice Hall, 2002, p. 215. ISBN 978-0-13-093315-7.
- ↑ Taiz, Lincoln. Plant Physiology and Development. Sunderland, MA: Sinauer Associates, Inc, 2015, p. 101. ISBN 978-1-60535-255-8.
- ↑ Freeman, Scott. Biological Sciences. United States of America: Pearson, 2014, p. 765–766. ISBN 978-0-321-74367-1.
- ↑ Simon, E.J., Dickey, J.L, & Reece, J.B. (2019). Campbell essential biology. 7th New York: Pearson
- ↑ Graham, Linda E. Plant Biology. Upper Saddle River, NJ 07458: Pearson Education, Inc., 2006, p. 200–202. ISBN 978-0-13-146906-8.
- ↑ «Capillary Action – Liquid, Water, Force, and Surface – JRank Articles». Science.jrank.org. Arxivat de l'original el 2013-05-27. [Consulta: 18 juny 2013].
- ↑ Mellander, Per-Erik; Bishop, Kevin; Lundmark, Tomas (en anglès) Forest Ecology and Management, 195, 1, 28-06-2004, pàg. 15-28. DOI: 10.1016/j.foreco.2004.02.051. ISSN: 0378-1127.
- ↑ Martin, John Holmes. Principles of Field Crop Production (en anglès). Macmillan, 1976. ISBN 978-0-02-376750-0.
- ↑ Jasechko, Scott; Sharp, Zachary D.; Gibson, John J.; Birks, S. Jean; Yi, Yi Nature, 496, 7445, 03-04-2013, pàg. 347–50. Bibcode: 2013Natur.496..347J. DOI: 10.1038/nature11983. PMID: 23552893.
- ↑ Evaristo, Jaivime; Jasechko, Scott; McDonnell, Jeffrey J. Nature, 525, 7567, 03-09-2015, pàg. 91–94. Bibcode: 2015Natur.525...91E. DOI: 10.1038/nature14983. ISSN: 0028-0836. PMID: 26333467.
- ↑ Bowen, Gabriel Nature, 525, 7567, 03-09-2015, pàg. 43–44. Bibcode: 2015Natur.525...43B. DOI: 10.1038/525043a. ISSN: 0028-0836. PMID: 26333464.
- ↑ Zhang, Yong-Jiang Plant Physiology, 172, 4, 12-2016, pàg. 2261–2274. DOI: 10.1104/pp.16.01191. PMC: 5129713. PMID: 27733514.
- ↑ Hochberg, Uri Plant Physiology, 174, 2, 6-2017, pàg. 764–775. DOI: 10.1104/pp.16.01816. PMC: 5462014. PMID: 28351909.
- ↑ 15,0 15,1 Holbrook, N. M.; Ahrens, E. T.; Burns, M. J.; Zwieniecki, M. A. «In vivo observation of cavitation and embolism repair using magnetic resonance imaging». Plant Physiology, 126, 1, 5-2001, pàg. 27–31. DOI: 10.1104/pp.126.1.27. ISSN: 0032-0889. PMC: 1540104. PMID: 11351066.
- ↑ Tiaz, Lincoln. Plant Physiology and Development. Massachusetts: Sinauer Associates, Inc., 2015, p. 63. ISBN 978-1605352558.
Bibliografia
[modifica]- Graham, Linda E.; Graham, Jim; Wilcox, Lee Warren. Plant Biology. 2a. Pearson Education, 2013, p. 680. ISBN 9781292042497.
- Nobel, Park S. Physicochemical and Environmental Plant Physiology. Academic Press, 2009, p. 582. ISBN 9780123741431.
- Salisbury, Frank B,; Ross, Cleon W. Plant Physiology. 4a. Wadsworth Publishing, 1992, p. 582. ISBN 9780534151621.
- Taiz, Lincoln; Zeiger, Eduardo. Plant Physiology. 5a. Sinauer, 2010, p. 782. ISBN 9780878935659.
Vegeu també
[modifica]Enllaços externs
[modifica]- USGS The Water Cycle: Evapotranspiration (water.usgs.gov)
| Disciplines de la botànica | .svg) | |
|---|---|---|
| Planta | ||
| Parts de les plantes | ||
| Cèl·lula vegetal | ||
| Reproducció vegetal | ||
| Taxonomia vegetal | ||
| Glossaris | ||
| ||