Alveolata

| Evolúciós időszak: Ediakara – Recens | ||||||||||
 | ||||||||||
| Rendszertani besorolás | ||||||||||
| ||||||||||
| Szinonimák | ||||||||||
| ||||||||||
| Kládok | ||||||||||
| Hivatkozások | ||||||||||
A Wikimédia Commons tartalmaz Alveolata témájú kategóriát. |
Az Alveolata (jelentése: „méhsejtszerűen üreges”)[1] egy főtörzsnek[2] tekintett eukariótaklád.[3] A sárgásmoszatokkal és a Rhizariával együtt a tubulokrisztás mitokondriumú protiszták Sar szupercsoportját alkotja.
Jellemzők
[szerkesztés]A legfőbb közös jellemzőik a kortikális (felszínközeli) alveolusok (zsákok). Ezek lapos vezikulumok egy réteggel az általuk támasztott sejtmembrán alatt, amelyek általában hajlékony pelliculus részei. A páncélos Dinoflagellata-tagokban ezek tartalmazhatnak kemény lemezeket. Az Alveolata tagjainak csőszerű bemélyedéseik vannak, és a sejteknek gyakran pórusszerű benyomódásaik vannak a felszínükön. A csoport szabadon élő és parazita szervezeteket, ragadozó ostorosokat és fotoszintetikus szervezeteket is tartalmaz.

Majdnem minden szekvenált csillós- és Apicomplexa-mitokondriálisgenom lineáris.[4] Majdnem minden mitokondrium saját mtDNS-t tartalmaz, de jelentősen csökkent genommal. Kivételek a csak mitoszómával rendelkező Cryptosporidium, a csillósok, és 2013-ban Janouškovec és társai kimutatták, hogy az Acavomonas korán elvált, így megőrzött némi génkódoló mtDNS-t.[5] A Babesia microti mitokondriális genomja körkörös.[6] Erről a fajról kiderült azonban, hogy se a Babesia, se a Theileria génuszba nem tartozik, így új génuszt kell neki létrehozni.
Történet
[szerkesztés]Az Apicomplexa, Dinoflagellata és csillósok kapcsolatát az 1980-as években feltételezték, és az 1990-es évek elején riboszomális RNS-ek összehasonlításával megerősítették, különösképpen Gajadhar és társai.[7] Cavalier-Smith vezette be az Alveolata nevet 1991-ben,[8] de ekkor parafiletikus csoportnak tekintette.[9]
Besorolás
[szerkesztés]Az Alveolata mintegy kilenc fő és kis csoportot tartalmaz. Ezek formái sokfélék, és ismert a kapcsolatuk ultrastrukturális és genetikai hasonlóságok révén:[10]
- Csillósok – gyakori protozoonok sok rövid csillóból álló sorokkal és két sejtmaggal
- Acavomonidia[10]
- Colponemidia[10]
- Dinoflagellata s.l. – többnyire tengeri ostorosok, sokuknak kloroplasztiszuk is van
- Perkinsozoa
- Chromerida – tengeri fotoszintetikus protozoonokból álló törzs
- Colpodellida
- Voromonadida
- Apicomplexa – parazita és másodlagosan nem fotoszintetikus protozoonok helyváltoztató mozgást végző axonémás szerkezetek nélkül, kivéve a gaméták esetén
Az Acavomonidia és a Colponemidia korábban kolponemidákként együtt szerepeltek, ezeket azonban különválasztották, mivel eltérő szerveződésük, illetve ultrastruktúrás azonosságuk van. Az Acavomonidia közelebb van a Dinoflagellata/Perkinsozoa csoporthoz a Colponemidiánál.[10] Így a kolponemida elnevezés két nem testvér csoportot jelent az Alveolatán belül: az Acavomonidiát és a Colponemidiát.[10]
Az Apicomplexa és a Dinoflagellata egymáshoz sokkal közelebbi kapcsolatban lehetnek a csillósoknál. Mindketteőnek van plasztiszuk, és azonosak a mikrotubulusokból álló szerkezetek a sejt tetején. Az Apicomplexa tagjaiban ez egy gazdasejtekbe kerüléshez használt komplex részét képezi, míg néhány színtelen Dinoflagellata-tagban ez egy pedunkulust képez, melyet zsákmányszerzéshez használ. Bizonyos további génuszok közeli kapcsolatban vannak e két csoporttal, melyek jellemzően hasonló csúcsszerkezetű ostorosok. Ide tartoznak szabadon élő Oxyrrhis- és Colponema-tagok, valamint parazita Perkinsus-,[11] Parvilucifera-, Rastrimonas- és Ellobiopsida-tagok. 2001-ben a tengeri pikoplankton minták rRNS-génjeinek másolása két új Alveolata-beli csoport jelenlétét bizonyította, melyeket I. és II. csoportnak neveztek el.[12][13] Az I. csoportnak nincs tenylsztett rokona, az II. a Dinoflagellata csoport Amoebophrya nevű parazitájának rokona, melyet korábban a Syndiniales rendbe soroltak.
Egyes tanulmányok szerint az Ascetosporea, mely főként tengeri gerinctelenek parazitáit tartalmazza, szintén ide tartozik, de nincsenek alveolusai, így az Endomyxa része.
Az Ellobiopsida kapcsolatai az Alveolatán belül ismeretlenek. Silberman és társai 2004-es tanulmánya filogenetikai analízis alapján a Thalassomyces génuszt, mely az Ellobiopsida része, az Alveolatába sorolta filogenetikai elemzéssel, azonban 2016-ban nem volt ismert bizonyosan a helyük.[14][15]
Filogenetika
[szerkesztés]2017-ben Thomas Cavalier-Smith az Alveolata filogenetikáját az alábbi módon írta le:[16]
| Alveolata |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||
Taxonómia
[szerkesztés]Alveolata Cavalier-Smith 1991 [Alveolatobiontes]
- Ciliophora törzs Doflein 1901 stat. n. Copeland 1956 [Ciliata Perty 1852; Infusoria Bütschli 1887; Ciliae, Ciliozoa, Cytoidea, Eozoa, Heterocaryota, Heterokaryota]
- Postciliodesmatophora altörzs Gerassimova & Seravin 1976
- Heterotrichea osztály Stein 1859
- Karyorelictea osztály Corliss 1974
- Intramacronucleata altörzs Lynn 1996
- ?Mesodiniea osztály Chen et al. 2015
- Lamellicorticata altörzság
- Litostomatea osztály Small & Lynn 1981
- Armophorea osztály Lynn 2004
- Cariacotrichea osztály Orsi et al. 2011
- Spirotrichea osztály Bütschli 1889
- Ventrata altörzság Cavalier-Smith 2004 [Conthreep Lynn 2012]
- ?Discotrichida rend Chen et al. 2015
- Protocruziea osztály Chen et al. 2015 [Protocruziidia de Puytorac, Grain & Mignot 1987]
- Colpodea osztály Small & Lynn 1981
- Nassophorea osztály Small & Lynn 1981
- Phyllopharyngea osztály de Puytorac et al. 1974
- Prostomatea osztály Schewiakoff 1896
- Plagiopylea osztály Small & Lynn 1985 sensu Lynn 2008
- Oligohymenophorea de Puytorac et al. 1974
- Postciliodesmatophora altörzs Gerassimova & Seravin 1976
- Miozoa törzs Cavalier-Smith 1987
- Colponemidia altörzs Tikhonenkov, Mylnikov & Keeling 2013
- Colponemea osztály Cavalier-Smith 1993
- Acavomonadia altörzs Tikhonenkov et al. 2014
- Acavomonadea osztály Tikhonenkov et al. 2014
- Subphylum Myzozoa Cavalier-Smith 2004
- Infraphylum Apicomplexa Levine 1970 emend. Adl et al. 2005
- Order ?Vitrellida Cavalier-Smith 2017
- ?Myzomonadea osztály Cavalier-Smith & Chao 2004 sensu Ruggiero et al. 2015
- Chromerea osztály
- Colpodellida rend Patterson & Zölffel 1991 [Spiromonadida Krylov & Mylnikov 1986]
- Sporozoa főosztály Leuckart 1879 stat. nov. Cavalier-Smith 2013 [Gamontozoa]
- Blastogregarinida osztály Chatton & Villeneuve 1936 [Blastogregarinina; Blastogregarinorina Chatton & Villeneuve 1936]
- Paragregarea osztály Cavalier-Smith 2014
- Gregarinomorphea osztály Grassé 1953
- Coccidiomorphea osztály Doflein 1901
- Dinozoa altörzság Cavalier-Smith 1981 emend. 2003
- ?Acrocoelida rend Cavalier-Smith & Chao 2004
- ?Rastromonadida rend Cavalier-Smith & Chao 2004
- Squirmidea osztály Norén 1999 stat. nov. Cavalier-Smith 2014
- Perkinsozoa főosztály Norén et al. 1999 s.s.
- Perkinsea osztály Levine 1978 [Perkinsasida Levine 1978]
- Dinoflagellata főosztály Butschli 1885 stat. nov. Cavalier-Smith 1999 sensu Cavalier-Smith 2013 [Dinozoa Cavalier-Smith 1981]
- Pronoctilucea osztály
- Ellobiopsea osztály Cavalier-Smith 1993 [Ellobiophyceae Loeblich III 1970; Ellobiopsida Whisler 1990]
- Myzodinea osztály Cavalier-Smith 2017
- Oxyrrhea osztály Cavalier-Smith 1987
- Syndinea osztály Chatton 1920 s.l. [Syndiniophyceae Loeblich III 1970 s.s.; Syndina Cavalier-Smith]
- Endodinea osztály Cavalier-Smith 2017
- Noctiluciphyceae osztály Fensome et al. 1993 [Noctilucae Haeckel 1866; Noctilucea Haeckel 1866 stat. nov.; Cystoflagellata Haeckel 1873 stat. nov. Butschli 1887]
- Dinophyceae osztály Pascher 1914 [Peridinea Ehrenberg 1830 stat. nov. Wettstein]
- Infraphylum Apicomplexa Levine 1970 emend. Adl et al. 2005
- Colponemidia altörzs Tikhonenkov, Mylnikov & Keeling 2013
Fejlődés
[szerkesztés]Az Alveolata plasztiszfejlődése érdekes. Cavalier-Smith szerint az Alveolata kloroplasztisszal rendelkező ősből fejlődött ki, mely a Chromista csoportot is létrehozta (Chromalveolata-hipotézis). Más kutatók szerint eredetileg az Alveolata tagjaiban nem voltak plasztiszok, és a Dinoflagellata és Apicomplexa külön szerezték meg. Azonban 2010-ben új elmélettel álltak elő, mely szerint az Alveolata, a Dinoflagellata, a Chromerida és a heterokont moszatok a plasztiszaikat egy vörösmoszatból szerezték, tehát ezen organellum eredete mind a négy csoportban közös.[17]
Evolúció
[szerkesztés]Egy Bayes-becslés szerint az Alveolata körülbelül 850 millió évvel ezelőtt fejlődhetett ki.[18] Az Alveolata tagjai a Myzozoa, a csillósok és a Colponemida. A Myzozoa kifejezés, melynek jelentése „a zsákmány tartalmának elszívása”, nem hivatalosan használatban lehetett az Alveolata egy részének közös ősére, melybe se a csillósok, se a Colponemida nem tartoznak. Az algapredáció fontos az Alveolata evolúciójában, ugyanis új plasztiszok endoszimbiózisára adhat forrásokat. A Myzozoa tehát lehetőség az Alveolata-történet követésére.
Az Alveolata ősei fotoszintetizálhattak.[19] Az ős valószínűleg rendelkezett plasztisszal. A Chromerida, az Apicomplexa és a Dinoflagellata peridinin tagjai megőrizték a plasztiszt.[20] Feltételezhető az is, hogy a Chromerida, a Dinoflagellata és a sárgásmoszatok közös monofiletikus plasztiszelődökkel is rendelkezhettek, vagyis plasztiszaikat egy vörösmoszatból szerezhették,[17] ami valószínűsíti, hogy az Alveolata és a Heterokonta közös őse is fotoszintetizált.
Egyesek szerint a Dinoflagellata, Apicomplexa, Colpodella, Chromerida és Voromonas közös őse mizocita predátor volt két független ostorral, mikropórusokkal, trichocisztákkal, roptriákkal, mikronémákkal, poláris gyűrűvel és nyílt oldalú konoiddal.[21] While the common ancestor of alveolates may also have possessed some of these characteristics, it has been argued that Myzocytosis was not one of these characteristics, as ciliates ingest prey by a different mechanism.[10]
Vitatott a plasztiszokat körülvevő membránok száma az Apicomplexa és a Dinoflagellata bizonyos tagjai esetén, valamint a membránok eredete. Ez az ultraszerkezeti jellemző felhasználható szervezetek csoportosítására, és ha közös a jellemző, az jelentheti, hogy a törzseknek közös fotoszintetizáló ősük volt. Mivel az Apicomplexa-fajok négy, a peridinin Dinoflagellata-tagok három membránnal körülvett plasztisszal rendelkeznek, Petersen és társai[22] nem tudták kizárni, hogy a közös Stramenopila-Alveolata plasztisz többször is hasznosnak bizonyult az Alveolatában – amelynek a forrásai Stramenopila-Alveolata donorok – bekebelezés és endoszimbiózis útján keresztül.
A csillósok modellszervezetek, mivel ezeket tanulmányozták nagymértékben a legtovább minden Alveolata-csoport közül. Különösek az eukarióták között, mivel a szaporodásukban a mikronukleusz és a makronukleusz is szerepet játszik. Szaporodásukat könnyen tanulmányozható, és modelleukariótává tette. Mivel csak predátorok, és nincs fennmaradó plasztiszuk, fejlődésük megmutatja, hogyan lehet egyensúlyban a predáció és az autotrófia,[19] és hogy ezen egyensúly az egyik vagy a másik irányba elbillenhet új törzs mixotróf ősökből való létrejöttekor, mely az egyik képesség elvesztését okozza.
-

-
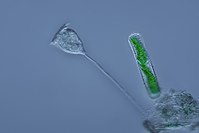 Vorticella (Ciliophora) (balra)
Vorticella (Ciliophora) (balra) -
 Plasmodium falciparum (Apicomplexa) a vérben
Plasmodium falciparum (Apicomplexa) a vérben -
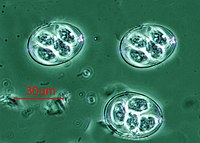 Eimeria maxima (Apicomplexa)
Eimeria maxima (Apicomplexa) -






Jegyzetek
[szerkesztés]- ↑ alveolate. Memidex (WordNet) Dictionary/Thesaurus. [2016. április 11-i dátummal az eredetiből archiválva]. (Hozzáférés: 2011. január 26.)
- ↑ Ruggiero MA, Gordon DP, Orrell TM, Bailly N, Bourgoin T, Brusca RC, Cavalier-Smith T, Guiry MD, Kirk PM (2015). „A higher level classification of all living organisms”. PLOS ONE 10 (4), e0119248. o. DOI:10.1371/journal.pone.0119248. PMID 25923521. PMC 4418965.
- ↑ Adl, S. M. (2012). „The revised classification of eukaryotes”. Journal of Eukaryotic Microbiology 59 (5), 429–514. o. DOI:10.1111/j.1550-7408.2012.00644.x. PMID 23020233. PMC 3483872.
- ↑ (2011) „The mitochondrial genome sequence of the ciliate Paramecium caudatum reveals a shift in nucleotide composition and codon usage within the genus Paramecium”. BMC Genomics 12, 272. o. DOI:10.1186/1471-2164-12-272. PMID 21627782. PMC 3118789.
- ↑ (2015. október 15.) „The Organellar Genomes of Chromera and Vitrella, the Phototrophic Relatives of Apicomplexan Parasites”. Annual Review of Microbiology 69 (1), 129–144. o, Kiadó: Annual Reviews. DOI:10.1146/annurev-micro-091014-104449. ISSN 0066-4227. PMID 26092225.
- ↑ (2012) „Sequencing of the smallest Apicomplexan genome from the human pathogen Babesia microti”. Nucleic Acids Res. 40 (18), 9102–14. o. DOI:10.1093/nar/gks700. PMID 22833609. PMC 3467087.
- ↑ Gajadhar, A. A. (1991). „Ribosomal RNA sequences of Sarcocystis muris, Theilera annulata, and Crypthecodinium cohnii reveal evolutionary relationships among apicomplexans, dinoflagellates, and ciliates”. Molecular and Biochemical Parasitology 45 (1), 147–153. o. DOI:10.1016/0166-6851(91)90036-6. PMID 1904987..
- ↑ Cavalier-Smith, T.. Cell diversification in heterotrophic flagellates, The Biology of free-living heterotrophic flagellates. Oxford University Press, 113–131. o. (1991). ISBN 978-0-19-857747-8
- ↑ Kumar, S. & Rzhetsky, A. 1996. Evolutionary relationships of eukaryotic kingdoms. Journal of Molecular Evolution, 42: 183–193
- ↑ a b c d e f Tikhonenkov, DV (2014). „Description of Colponema vietnamica sp.n. and Acavomonas peruviana n. gen. n. sp., two new alveolate phyla (Colponemidia nom. nov. and Acavomonidia nom. nov.) and their contributions to reconstructing the ancestral state of alveolates and eukaryotes”. PLOS ONE 9 (4), e95467. o. DOI:10.1371/journal.pone.0095467. PMID 24740116. PMC 3989336.
- ↑ (2011) „Spliced leader RNAs, mitochondrial gene frameshifts and multi-protein phylogeny expand support for the genus Perkinsus as a unique group of Alveolates”. PLOS ONE 6 (5), e19933. o. DOI:10.1371/journal.pone.0019933. PMID 21629701. PMC 3101222.
- ↑ (2001) „Unexpected diversity of small eukaryotes in deep-sea Antarctic plankton”. Nature 409 (6820), 603–7. o. DOI:10.1038/35054537. PMID 11214316.
- ↑ (2001) „Oceanic 18S rDNA sequences from picoplankton reveal unsuspected eukaryotic diversity”. Nature 409 (6820), 607–10. o. DOI:10.1038/35054541. PMID 11214317.
- ↑ Hoppenrath, Mona (2016. április 29.). „Dinoflagellate taxonomy — a review and proposal of a revised classification”. Marine Biodiversity 47 (2), 381–403. o, Kiadó: Senckenberg Institute (Springer). DOI:10.1007/s12526-016-0471-8. ISSN 1867-1616.
- ↑ Taylor, F. J. R. "Max" (2004). „Illumination or confusion? Dinoflagellate molecular phylogenetic data viewed from a primarily morphological standpoint”. Phycological Research 52 (4), 308–324. o, Kiadó: Japanese Society of Phycology (Wiley). DOI:10.1111/j.1440-183.2004.00360.x. ISSN 1322-0829.
- ↑ Cavalier-Smith, Thomas (2017. szeptember 5.). „Kingdom Chromista and its eight phyla: a new synthesis emphasising periplastid protein targeting, cytoskeletal and periplastid evolution, and ancient divergences”. Protoplasma 255 (1), 297–357. o. DOI:10.1007/s00709-017-1147-3. PMID 28875267. PMC 5756292.
- ↑ a b (2010) „A common red algal origin of the apicomplexan, dinoflagellate, and heterokont plastids”. Proc Natl Acad Sci USA 107 (24), 10949–54. o. DOI:10.1073/pnas.1003335107. PMID 20534454. PMC 2890776.
- ↑ (2006) „A molecular time-scale for eukaryote evolution recalibrated with the continuous microfossil record”. Proc Biol Sci 273 (1596), 1867–72. o. DOI:10.1098/rspb.2006.3537. PMID 16822745. PMC 1634798.
- ↑ a b Reyes-Prieto, A (2008). „Multiple genes of apparent algal origin suggest ciliates may once have been photosynthetic”. Curr. Biol. 18 (13), 956–962. o. DOI:10.1016/j.cub.2008.05.042. PMID 18595706. PMC 2577054.
- ↑ Moore RB, Oborník M, Janouskovec J, Chrudimský T, Vancová M, Green DH, Wright SW, Davies NW, Bolch CJ, Heimann K, Slapeta J, Hoegh-Guldberg O, Logsdon JM, Carter DA (2008). „A photosynthetic alveolate closely related to apicomplexan parasites”. Nature 451 (7181), 959–963. o. DOI:10.1038/nature06635. PMID 18288187.
- ↑ (2002) „The phylogeny of colpodellids (Alveolata) using small subunit rRNA gene sequences suggests they are the free living sister group to apicomplexans”. J Eukaryot Microbiol 49 (6), 498–504. o. DOI:10.1111/j.1550-7408.2002.tb00235.x. PMID 12503687.
- ↑ Petersen J, Ludewig AK, Michael V, Bunk B, Jarek M, Baurain D, Brinkmann H (2014). „Chromera velia, endosymbioses and the rhodoplex hypothesis—plastid evolution in cryptophytes, alveolates, stramenopiles, and haptophytes (CASH lineages)”. Genome Biol Evol 6 (3), 666–684. o. DOI:10.1093/gbe/evu043. PMID 24572015. PMC 3971594.