Сестринский хроматидный обмен

Сестринский хроматидный обмен (СХО) — это обмен участками между сестринскими хроматидами одной хромосомы. Процесс обмена происходит во время S-фазы клеточного цикла путём гомологичной рекомбинации между сестринским хроматидами, генетическая информация при этом остаётся неизменной.
Сестринские хроматидные обмены выявляют в метафазах второго клеточного деления после добавления к пролиферирующим клеткам модифицированных нуклеозидов, способных после фосфорилирования встраиваться в ДНК при репликации. В качестве таких нуклеозидов используют, как правило, аналоги тимидина, такие как 5-бромдезоксиуридин или 5-этинилдезоксиуридин.
Впервые сестринские хроматидные обмены увидел американский генетик Джеймс Г. Тейлор (англ. J. Herbert Taylor) в 1958 году в апикальных клетках корня лилийного растения Bellevalia romana[1]. В качестве модифицированного аналога тимидина он использовал тимидин, меченый тритием[2].
Сестринские хроматидные обмены происходят спонтанно, их средняя частота в лимфоцитах человека в норме составляет 3—4 на клетку. Повышение частоты сестринских хроматидных обменов происходит при воздействии факторов, вызывающих репликативный стресс, поперечные сшивки ДНК или блокировку репликационной вилки[3]. Повышенная частота сестринских хроматидных обменов обнаружена в клетках больных синдромом Блума[4].
-
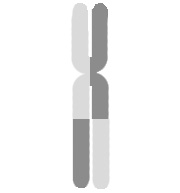 Схематическое изображение сестринского хроматидного обмена
Схематическое изображение сестринского хроматидного обмена -
 Сестринские хроматидные обмены в клетках HeLa
Сестринские хроматидные обмены в клетках HeLa -
 Сестринские хроматидные обмены в ЭСК человека.
Сестринские хроматидные обмены в ЭСК человека.



Дифференциальная окраска сестринских хроматид
[править | править код]Для получения дифференциальной окраски хроматид клетки культивируют in vitro в присутствии бромдезоксиуридина в течение двух клеточных циклов. В результате этого получаются хромосомы, у которых тимидин в одной хроматиде замещён бромдезоксиуридином в обеих цепях ДНК, а в сестринской хроматиде — только в одной из цепей. После этого препарат метафазных хромосом, полученный из таких клеток, красят фотосенсибилизирующим флуоресцентным красителем Hoechst33258 и облучают ультрафиолетовым светом, вследствие чего происходит фотодеградация ДНК[5]. Хроматида, в которой тимидин замещен в обоих цепях ДНК, становится неспособной связывать краситель Гимза и остаётся при окраске бледной, а хроматида, в которой бромдезоксиуридин присутствует только в одной цепи ДНК, способна окрашиваться как обычный хроматин и станет тёмной при окраске. Полученную окраску называют дифференциальной окраской сестринских хроматид[1]. Дифференциальную окраску сестринских хроматид иногда называют арлекиновой окраской.
Примечания
[править | править код]- ↑ 1 2 Коряков Д. Е., Жимулев И. Ф. Хромосомы. Структура и функции. — Новосибирск: Из-во СО РАН, 2009. — 258 с. — ISBN 978-5-7692-1045-7.
- ↑ Taylor J. H. Sister Chromatid Exchanges in Tritium-Labeled Chromosomes (англ.) // Genetics : journal. — 1958. — May (vol. 43, no. 3). — P. 515—529. — PMID 17247775. — PMC 1209899. Архивировано 5 июля 2022 года.
- ↑ David M. Wilson III and Larry H. Thompson: Molecular mechanisms of sister-chromatid exchange. Mutat. Res. 2007; 616(1-2):11—23. PMID 17157333.
- ↑ German J, Schonberg S, Louie, and Chaganti R S. Bloom’s syndrome. IV. Sister-chromatid exchanges in lymphocytes. Am. J. Hum. Genet. 1977; 29(3): 248—255. PMID 868871.
- ↑ Goto K., Akematsu T., Shimazu H., Sugiyama T. Simple differential Giemsa staining of sister chromatids after treatment with photosensitive dyes and exposure to light and the mechanism of staining. Chromosoma, 1975; 53(3):223—230. PMID 53133.
Литература
[править | править код]- Захаров А. Ф., Бенюш В. А., Кулешов Н. П., Барановская Л. И. Хромосомы человека. Атлас. — М.: Медицина, 1982. — 263 с.
- Коряков Д. Е., Жимулев И. Ф. Хромосомы. Структура и функции. — Новосибирск: Из-во СО РАН, 2009. — 258 с. — ISBN 978-5-7692-1045-7.
- Mezzanotte, R., and Nieddu, M. A Historical Overview of Bromo-Substituted DNA and Sister Chromatid Differentiation // Functional Analysis of DNA and Chromatin / J. C. Stockert, J. Espada, and A. Blázquez-Castro. — Totowa, NJ: Humana Press, 2014. — С. 88—89. — (Methods in Molecular Biology). — ISBN 978-1-62703-705-1.
| Основное |  | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Классификация | |||||||||||
| Структура |
| ||||||||||
| Перестройки и нарушения | |||||||||||
| Хромосомное определение пола | |||||||||||
| Методы | |||||||||||