Salida de las aguas


En la historia evolutiva de los seres vivos, la salida de las aguas, también llamada conquista de los continentes, conquista de las tierras o terrestrificación (término preferible),[2] corresponde a la adaptación progresiva, durante el Arcaico por las bacterias y durante el Paleozoico, por la flora y luego la fauna, hasta aquí esencialmente marinas, a un modo de vida terrestre. Las extinciones del Ordovícico-Silúrico y la del Devónico estuvieron marcadas notablemente por importantes crisis biológicas que empobrecieron la vida marina y favorecieron la conquista de las tierras emergidas por las plantas clorofílicas y varios grandes grupos de animales, principalmente los moluscos, los artrópodos y los vertebrados Esta conquista por las primeras formas vegetales habría tenido lugar en el Cámbrico hace alrededor de 500 millones de años, por las primeras plantas vasculares en el límite Ordovícico-Silúrico (445 millones de años),[3] y por las formas animales terrestres —artrópodos, vertebrados entre otros— alrededor de 430 millones de años.[4] Muchas adaptaciones se desarrollaron, tanto por los vegetales como por los animales, el paso de la vida acuática a la vida terrestre y a la respiración aérea representan un salto macroevolutivo importante en la historia de los organismos vivos.[5] De hecho, este proceso se repite y ocurre independientemente durante la historia de los seres vivos, ya sea por los organismos unicelulares, los hongos, las plantas, los invertebrados o los vertebrados. Por lo tanto, no se debería hablar de una sino de varias salidas de las aguas.[6]
Finalmente, este mito de la salida única del agua tiende a enmascarar el hecho de que ha habido muchos «retornos al agua». Entre los vertebrados tetrápodos que han regresado al agua secundariamente se encuentran las tortugas, ictiosaurios, los plesiosaurios, los mosasaurios, los cocodrilos, algunos escamosos (serpientes marinas, varias iguanas) y cuatro líneas de mamíferos (cetáceos, sirenias, Pinnípedos y Lutrinae).[7]
Historia evolutiva[editar]
Salida de las aguas de las bacterias[editar]
Los principales grupos de fotótrofos por el carbono, procariotas o eucariotas, aparecieron en el medio marino. Sin embargo, un estudio realizado en 2017 sugiere que las bacterias incursionaron en las aguas termales de agua dulce terrestre hace 3500 millones de años.[8]
Aun así, existen argumentos geoquímicos indirectos y rastros fósiles directos de biopelículas bacterianas a favor de la aparición de diferentes bacterias en la Tierra varias veces durante el Arcaico y la de las cianobacterias hace 1000 millones de años.[9] También es probable que se produzcan microhongos acuáticos en el fondo del océano en respiraderos hidrotermales, y la hidromicoflora ha mantenido desde entonces un estilo de vida bentónico.[10][11]
El establecimiento de la simbiosis en un entorno acuoso ha permitido proporcionar soluciones y dar un salto macroevolutivo para la conquista del medio terrestre más hostil por parte de las plantas: diversas asociaciones mutualistas (líquenes, micophycobiosis como Stigmidium ascophylli en el Cámbrico, la micorrización por glomeromicetos silúricos[12]) con los fotótrofos se utilizaron simultánea o sucesivamente.[13][14] Los investigadores suponen que las primeras comunidades terrestres formaron costras en los suelos, recorridas progresivamente por artrópodos.[15]
Salida de las aguas de la flora y después de la fauna[editar]




Después de la aparición de las algas en los océanos, la falta de agua y las fuertes radiaciones ultravioletas sobre la tierra habrían confinado a esas algas ancestrales en ambientes acuáticos. La evolución de las innovaciones relativas a la reproducción (ciclo de vida dominado por la fase esporofítica), a la arquitectura (planes corporales) y a la protección contra las pérdidas de agua habría allanado el camino para la adaptación de las plantas a la tierra firme.[19]
El Ordovícico vio una explosión de la biodiversidad conocida como la gran biodiversidad ordovícica. La vida en ese momento estaba esencialmente confinada a los mares y a los océanos, especialmente en los mares epicontinentales poco profundos que bordeaban los muchos continentes, con aguas poco profundas que permitían en ellos la fotosíntesis. Las altas productividades diarias y anuales de fitoplancton se encuentran entonces en esos ecosistemas costeros y correspondían a la máxima biodiversidad del medio marino, la abundancia de fitoplancton afectaba al resto de la cadena alimentaria hasta a los peces con mandíbula.[20]
Los embriofitos (es decir, las plantas terrestres) que aparecieron hace unos 450 millones de años probablemente derivan de las algas verdes del grupo de los carofitass, que evolucionan en agua dulce y fueron capaces de resistir emersiones temporales gracias a esas asociaciones simbióticas.[21] La conquista terrestre requirió varias adaptaciones simultáneas, estas algas probablemente adoptaron una estrategia simbiótica (micophycobioses, líquenes[22]) para realizar la primera salida de las aguas de las plantas antes que la de las plantas vasculares (del tipo polisporangiadas asociado con micorrizas[23]). La conquista terrestre de hecho requirió varias adaptaciones simultáneas para responder a las tensiones engendradas por ese entorno:[24]
- baja disponibilidad de agua con variaciones muy grandes, con fases de ausencia, que resultan en una alimentación mineral irregular; recursos nutritivos regionalizados (agua y sales minerales en el sustrato, gas y luz en el aire). La forma filamentosa y la producción de exoenzimas de hongos explotan mejor estos recursos edáficos, mientras que la ramificación de los talos clorofílicos garantiza la explotación de los recursos en el aire.
- luz atmosférica más rica en radiación, especialmente en ultravioleta, que la luz filtrada por el agua, lo que requiere el desarrollo de mecanismos fotoprotectores (pigmentos fúngicos y luego cutícula):
- el ambiente está mucho menos protegido térmicamente que el mar (diferencias rápidas de temperatura y gran amplitud), de ahí el desarrollo de talos más grandes que el parénquima que aseguran la homeohidria;
- empuje de Arquímedes reducido en el medio aéreo, causando problemas de sustento, lo que impone formas vegetales simples y después complejas a través del desarrollo de un metabolismo fenólico que permite la síntesis de biopolímeros esenciales, la cutina, la celulosa y la lignina que aseguran una impermeabilización. de la planta, una protección contra las radiaciones y la rigidización de sus partes erigidas.[25]
El desarrollo de estos embriofitos se vio favorecido por los hongos que, al secretar enzimas exocelulares, tenían la capacidad de disolver los sustratos minerales del suelo.[15]
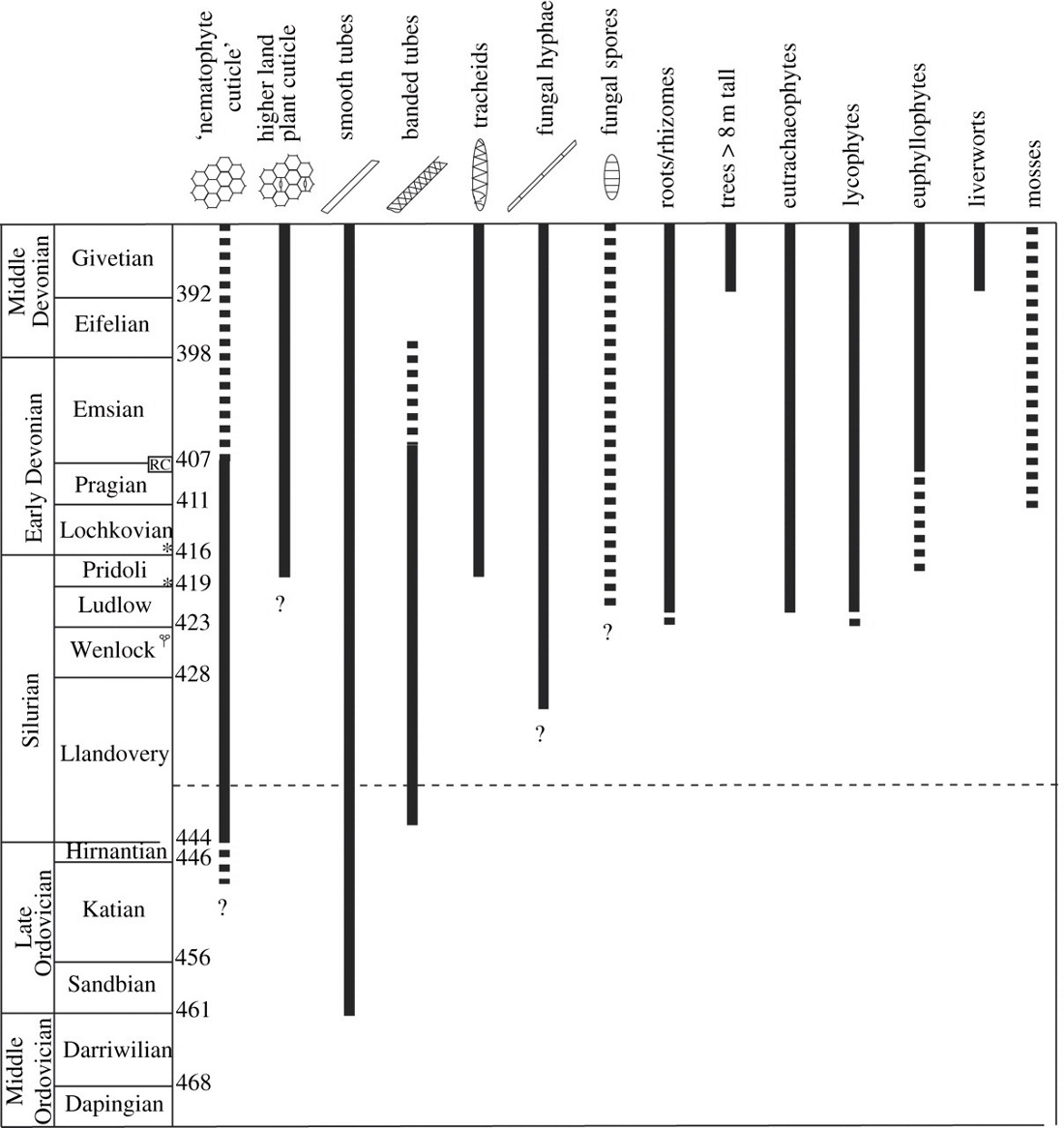
El registro fósil de los primeras vegetales terrestres consiste en gran parte en microfósiles (criptosporas y luego esporas triletadas[26]), que datan de alrededor de 460 Ma, y en desechos macroscópicos.[27] Las primeras plantas de las que se disponen restos casi completos pertenecen al taxón de las Rhyniopsida, que vivían en áreas (márgenes de mares, zonas de inundación) donde experimentaron emersiones ocasionales. La ausencia de raíces y de hojas limitaba su tamaño, sus «tallos»[28] eran ejes ramificados con ramas dicotómicas en las que se insertaban los esporangios. El desarrollo de los tejidos conductores, de los tallos y luego de los sistemas radiculares favoreció progresivamente el aumento del tamaño de las plantas vasculares en el ambiente aéreo.[29] Dos extinciones masivas ocurrieron al final de este período, entre 450 y 440 millones de años. Las extinciones del Ordovícico-Silúrico estuvieron marcada por una importante glaciación y una bajada del nivel del mar. Esta regresión marina vio una retirada del mar a lo largo de cientos de kilómetros, lo que empobreció la vida marina de las plataformas continentales. Otra consecuencia de esta regresión fue la expansión de los ambientes de agua dulce, ya que la escorrentía continental tenía que recorrer distancias más largas antes de llegar a los océanos. Esta extinción fue seguida por un evento anóxico oceánico vinculado a una gran transgresión marina global que también causó trastornos ecológicos que dificultaron la adaptación de las especies a sus ecosistemas marinos. Esto condujo en el Silúrico al desarrollo de los Eutracheophytes (verdaderas plantas vasculares) y de artrópodos (miriápodos del tipo Pneumodesmus newmani, arañas, ácaros, colémbolos, mientras que los anélidos probablemente hayan conquistado las tierras emergidas desde el Ordovícico) en ecosistemas terrestres marginales (marismas costeras, lagunas arenosas, riberas de ríos y lagos).
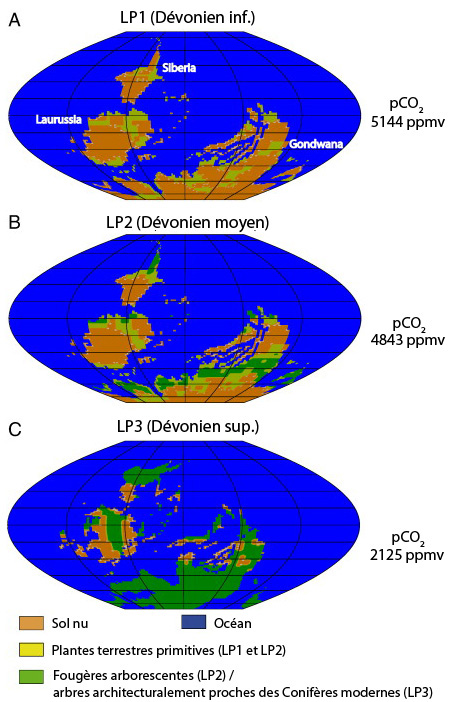
Aún subordinados a los humedales, las plantas terrestres (incluidas los traqueofitos), los moluscos (incluidos los gasterópodos), los artrópodos (incluidos los insectos) y los vertebrados emergerán gradualmente de las aguas para convertirse en verdaderos terrestres.[6] Durante el Devónico esta colonización progresiva de la tierra,[30] en varias etapas,[31] se vio favorecida por:
- el aumento de la tasa de oxígeno marino y atmosférico, que hasta entonces era un factor limitante: mientras que la atmósfera se enriquecía con oxígeno gracias al desarrollo de la fotosíntesis, O
2 sufría la acción de la radiación ultravioleta y se transformaba en parte en ozono. Así se formó gradualmente una capa de ozono, que desempeñó su papel de pantalla en el Silúrico, filtrando una gran parte de las radiaciones letales del Sol.[32] - la extensión de las planicies intermareales durante los períodos cálidos con un nivel general de los mares elevados, cuando en las plataformas continentales se desarrollaban formaciones de plantas de tipo «manglar» (pero consistentes en especies con esporas), favoreciendo a las especies animales con apéndices o aletas sólidas y cavidades o tubos respiratorios o incluso vejigas natatorias ricamente vascularizadas y plegadas, capaces de extraer oxígeno del aire O
2 durante la marea baja.[33]
La extinción del Devónico entre 408 y 360 millones de años (eustatismo, evento anóxico, cambios climáticos) afectó principalmente a los invertebrados (amonitas y trilobites) y vertebrados (Placodermos, Sarcopterigios) marinos y de agua dulce. Esto favoreció el desarrollo de los vertebrados modernos, que consisten principalmente en actinopterigios, condrictios y tetrápodos cuyas primeras salidas de agua conocidas datan del Alto Devónico, hacia 365 millones de años, con Ichthyostega, el más antiguo de los vertebrados conocidos adaptado tanto a la natación como a una forma de locomoción terrestre. La adaptación de los tetrápodos al medio terrestre se vuelve muy pronunciada durante el Carbonífero (-359 a 299 millones de años), lo que lleva a los tetrápodos actuales que son los anfibios, los reptiles, las aves y los mamíferos.[34] La reducción de la pérdida de agua en el ambiente del aire se vio favorecida por la permeabilidad drásticamente más baja de su tegumento en comparación con las especies acuáticas, la internalización de sus superficies respiratorias, una orina escasa y heces poco hidratadas, una alimentación sólida y líquida que permitió ganancias en agua y en sal. Finalmente, muchas adaptaciones conductuales complementaron las adaptaciones morfoanatómicas y fisiológicas, como la actividad nocturna.[35]
De los 31 filos de animales actuales identificados, todos los cuales aparecieron en el medio marino, 12 son endémicos de este medio, 19 han conquistado la tierra.[36]
¿Otra historia?[editar]
La extinción al final del Devónico afectó al 70% de las especies vivas y más particularmente a las especies marinas, pero afectó relativamente poco a los artrópodos que ya estaban muy diversificados y a los primeros vertebrados tetrápodos. Según el profesor de paleobiología George R. McGhee, si esta extinción masiva hubiera sido más severa, la historia de la vida en la Tierra habría sido completamente alterada. La larga marcha de los tetrapodomorfos que evolucionaron a partir de los peces sarcopterigios se habría detenido y la conquista de la tierra habría sido asegurada principalmente por los tres grupos principales de artrópodos actuales (los hexapodos —insectos y colémbolos—, los miriápodos —ciempiés— y los arácnidos —arañas, ácaros y escorpiones) —.[37] Una extinción aún más masiva habría llevado a un planeta microbiano. Además, la biodiversidad microbiana todavía representa el 80% de la biomasa total del planeta[38] y, según el profesor de microbiología Jean-Louis Fauchère, los hombres son «los únicos avatares del mundo bacteriano».[39] No es sorprendente que la terrestrificación de los tres grupos principales, los vegetales, los artrópodos y los vertebrados haya sido acompañada por el uso de las actividades de los microorganismos, que ya son la forma dominante del mundo vivo en términos de abundancia y biodiversidad desde la aparición de la vida en la Tierra. Las especies de estos tres grupos son comunidades simbióticas por su origen y han adoptado la misma estrategia común de servirse de mutualistas microbianos (bacterias, protistas y microhongos), tanto interna como externamente para desarrollar asociaciones mutualistas para las plantas terrestres (concepto de fitobioma y de microbiota de plantas, especialmente con micorrizas), de desintoxicar parcialmente la materia vegetal y aumentar considerablemente el valor calorífico y nutritivo disponible para los animales (noción de microbiota intestinal) o para protegerlos (microbiota oral), cutánea, vaginal...).[40][41]
Véase también[editar]
Notas[editar]
- ↑ Estos huesos llamados radiales, están dispuestos en tres filas transversales y se segmentan, proceso que podría estar en el origen de los metacarpianos y las falanges. Según N. H. Shubin & P. Alberch (1986). «A morphogenetic approach to the origin and basic organization of the tetrapod limb». Evolutionary Biology (en inglés) 20 (1): 319-387..
- ↑ Terrestrificación es un término menos antropomorfizante, el de conquista o franqueo sobrentiende un cierto derecho en relación al medio acuoso original y una cierta falla de los organismos que permanecieron en el agua. Esta expresión también corresponde a una visión finalista, como si los seres vivos tuvieran la voluntad de abandonar el agua, mientras que es el agua la que se retira por razones climáticas o geológicas. (Véase Sébastien Steyer, La Terre avant les dinosaures, Belin, 2009, p. 7 ). Además, la mayoría de los vertebrados son acuáticos: con 25 000 especies conocidas —100 descritas cada año— y aproximadamente el mismo número aún desconocido, los peces son el grupo más grande de vertebradoss ((en inglés) Véase: Quentin Bone, Richard Moore, Biology of Fishes, Taylor & Francis, 2008, p. 1 ). Finalmente, en términos de fisiología, este franqueo es muy incompleto, los animales y plantas terrestres no están completamente «salidos de las agua» con un cuerpo compuesto principalmente de agua. Sin embargo, el uso de expresiones antropomorfizantes por parte de los científicos puede ser útil según el botánico Lucien Baillaud quien escribe:
. Véase: Lucien Baillaud (2007). «La végétalité : un réseau de déterminismes reliant structure, biologie et milieu de vie dans le deuxième royaume de la nature». Acta Botanica Gallica 154 (2): 153-201..No desprecies el antropomorfismo si nos ayuda a expresarnosNe méprisons pas l'anthropomorphisme s'il nous aide à nous exprimer - ↑ Jennifer L. Morris, Mark N. Puttick, James W. Clark, Dianne Edwards, Paul Kenrick, Silvia Pressel, Charles H. Wellman, Ziheng Yang, Harald Schneider & Philip C. J. Donoghue (20 février 2018). «The timescale of early land plant evolution». PNAS (en inglés). doi:10.1073/pnas.1719588115..
- ↑ El descubrimiento en 2004 de un ciempiés, Pneumodesmus newmani, datado en 442 millones de años, corresponde a uno de los artrópodos terrestres más antiguos conocidos. Véase, Heather M. Wilson & Lyall I. Anderson (2004). «Morphology and taxonomy of Paleozoic millipedes (Diplopoda: Chilognatha: Archipolypoda) from Scotland». Journal of Paleontology (en inglés) 78 (1): 169-184. doi:10.1666/0022-3360(2004)078<0169:MATOPM>2.0.CO;2.
- ↑ Gilles Bœuf (mayo de 2011). «Marine biodiversity characteristics». Comptes Rendus Biologies (en inglés) 334 (5-6): 435-440. doi:10.1371/journal.pone.0066906..
- ↑ a b Du Silurien au Dévonien : les sorties des eaux, dossier CNRS
- ↑ Guillaume Lecointre (2014). [Salida de las aguas en Google Libros L'évolution, question d'actualité ?]
|url=incorrecta (ayuda). éditions Quae. p. 79. - ↑ Tara Djokic, Martin J. Van Kranendonk, Kathleen A. Campbell, Malcolm R. Walter & Colin R. Ward (2017). «Earliest signs of life on land preserved in ca. 3.5 Ga hot spring deposits». Nature Communications (en inglés) 8 (15263). doi:10.1038/ncomms15263..
- ↑ Fabia U. Battistuzzi S. Blair Hedges (février 2009). «A Major Clade of Prokaryotes with Ancient Adaptations to Life on Land». Molecular Biology and Evolution (en inglés) 26 (2): 335-343. doi:10.1093/molbev/msn247..
- ↑ Butterfield, N. J. (2005). Probable Proterozoic Fungi. Cambridge.
- ↑ Jean-Christophe Guéguen, David Garon (2015). Biodiversité et évolution du monde fongique. EDP Sciences. p. 92..
- ↑ Al mismo tiempo, se descubrieron los fósiles de Prototaxites cuya naturaleza fúngica se discute.
- ↑ Marc-André Selosse, François Le Tacon (2001). «Les stratégies symbiotiques de conquête du milieu terrestre par les végétaux». Année Biol. 40: 16-17. Archivado desde el original el 11 de noviembre de 2017. Consultado el 5 de octubre de 2019.
- ↑ Jean-Christophe Guéguen, David Garon (2015). Biodiversité et évolution du monde fongique. EDP Sciences. p. 89..
- ↑ a b Francis Martin (2019). Sous la forêt. Pour survivre il faut des alliés. Humensis. p. 121..
- ↑ Grzegorz Niedźwiedzki et al (7 janvier 2010). «Tetrapod trackways from the early Middle Devonian period of Poland». Nature (en inglés) (463): 43-48. doi:10.1038/nature08623..
- ↑ Con varios clados : Agnathans (lampreas), condrictios (tiburones, rayas), Placodermos (fósiles), Acanthodianos (fósiles), Osteichthyanos (peces óseos).
- ↑ « Systématique : ordonner la diversité du vivant »,' Rapport sur la Science et la technologie N°11, Académie des sciences, Lavoisier, 2010, p. 65
- ↑ Peter H Raven, Kenneth A Mason, Georges B Johnson, Jonathan B Losos, Susan R Singer (2017). [Salida de las aguas en Google Libros Biologie]
|url=incorrecta (ayuda). De Boeck Superieur. p. 588. - ↑ B. D. Webby, Florentin Paris, Mary L. Droser (2013). The Great Ordovician Biodiversification Event (en inglés). Columbia University Press. p. 496.
- ↑ Douglas E. Soltis, Pamela Soltis, J.J. Doyle (1998). Molecular Systematics of Plants II : DNA Sequencing (en inglés). Springer. p. 530.
- ↑ Por provocación, es tentador escribir que la mayoría de los archégoniates terrestres son tipos de líquenes, cuyo alga es multicelular y ocupa la parte aérea y visible de la asociaciónPar provocation, il est tentant d'écrire que la majorité des Archégoniates terrestres sont des sortes de lichens, dont l'algue est pluricellulaire et occupe la partie aérienne et visible de l'associationMarc-André Selosse, p. 16.
- ↑ Jean-Christophe Guéguen, David Garon (2014). Biodiversité et évolution du monde fongique. EDP Sciences. p. 95.
- ↑ Marc-André Selosse, François Le Tacon (2001). «Les stratégies symbiotiques de conquête du milieu terrestre par les végétaux». Année Biol. 40: 4.
- ↑ Hugues Renault, Annette Alber, Nelly A. Horst, Alexandra Basilio Lopes, Eric A. Fich, Lucie Kriegshauser, Gertrud Wiedemann, Pascaline Ullmann, Laurence Herrgott, Mathieu Erhardt, Emmanuelle Pineau, Jürgen Ehlting, Martine Schmitt, Jocelyn K. C. Rose, Ralf Reski & Danièle Werck-Reichhart (2017). «A phenol-enriched cuticle is ancestral to lignin evolution in land plants». Nature Communications (en inglés) 8 (14713). doi:10.1038/ncomms14713..
- ↑ Con tres fentas de dehiscencia.
- ↑ Registre fossile des types de fragments et des groupes taxonomiques de plantes terrestres de l'Ordovicien moyen au Dévonien moyen
- ↑ No se puede usar este término porque no tienen sus estructuras diferenciadas actuales.
- ↑ P. G Gensel (2008). «The Earliest Land Plants». Annual Review of Ecology, Evolution, and Systematics (en inglés) 39 (1): 459-477.
- ↑ Répartitions potentielles de la végétation à trois périodes du Dévonien, calculées par le modèle de G. Le Hir et al., tiré de (en inglés) Le Hir, G., Y. Donnadieu, Y. Goddéris, B. Meyer-Berthaud, G. Ramstein, R. C. Blakey, 2011. The climate change caused by the land plant invasion in the Devonian. Earth and Planetary Science Letters, 310(3–4): 203-212
- ↑ Sébastien Steyer (2008). La Terre avant les dinosaures. Belin ("Pour la Science"). p. 76-80. ISBN 9782701142067.
- ↑ Jean-Claude Roland, Hayat El Maarouf Bouteau, François Bouteau (2008). Atlas de biologie végétale. Dunod. p. 122.
- ↑ Patrick De Wever, Bruno David, Didier Néraudeau, Jean Broutin, Philippe Janvier et al. (2010). Paléobiosphère : regards croisés des sciences de la vie et de la terre. Vuibert, MNHN, SGF. p. 269-275. ISBN 9782711725038.
- ↑ Lauren Cole Sallan et Michael I. Coates (1 de junio de 2010). «End-Devonian extinction and a bottleneck in the early evolution of modern jawed vertebrates». Proceedings of the National Academy of Sciences (en inglés) 107 (22). doi:10.1073/pnas.0914000107.
- ↑ Denis Poinsot, Maxime Hervé, Bernard Le Garff, Mael Ceillier (2018). [Salida de las aguas en Google Libros Diversité animale. Histoire, évolution et biologie des Métazoaires]
|url=incorrecta (ayuda). De Boeck Superieur. p. 173.. - ↑ Gilles Bœuf (mayo de 2011). «Marine biodiversity characteristics». Comptes Rendus Biologies (en inglés) 334 (5-6): 435. doi:10.1016/j.crvi.2011.02.009..
- ↑ George R. McGhee (2013). When the invasion of land failed. The legacy of the devonian extinctions (en inglés). Columbia University Press. p. 61-67..
- ↑ William B. Whitman, David C. Coleman & William J. Wiebe (junio de 1998). «Prokaryotes: The unseen majority». PNAS (en inglés) 95 (12): 6578-6583. doi:10.1073/pnas.95.12.6578..
- ↑ Jean-Louis Fauchère (2017). Les Bactéries et l’Homme: Une cohabitation tumultueuse. Editions Edilivre. p. 137. «que des avatars du monde bactérien»..
- ↑ P. Engel, NA. Moran (septembre 2013). «The gut microbiota of insects - diversity in structure and function». FEMS Microbiol Rev. (en inglés) 37 (5): 699-735. doi:10.1111/1574-6976.12025..
- ↑ R.E. Ley, M. Hamady, C. Lozupone, P.J.Turnbaugh, R.R. Ramey, J.S. Bircher et al. (septembre 2008). «Evolution of mammals and their gut microbes». Science (en inglés) 320 (5883): 1647-1651. doi:10.1126/science.1155725..
{kind=link}
{kind=link}
Referencias[editar]
- Esta obra contiene una traducción derivada de «Sortie_des_eaux» de Wikipedia en francés, publicada por sus editores bajo la Licencia de documentación libre de GNU y la Licencia Creative Commons Atribución-CompartirIgual 4.0 Internacional.
Bibliografía[editar]
- Pierre Corvol, Jean-Luc Elghozi (2011). [Salida de las aguas en Google Libros Sortir de l’eau. De la vie aquatique à la vie]
|url=incorrecta (ayuda). Odile Jacob. p. 240. - George R. McGhee Jr. (2013). [Salida de las aguas en Google Libros When the invasion of land failed. The legacy of the Devonian extinctions]
|url=incorrecta (ayuda) (en inglés). Columbia University Press. p. 336.
Enlaces externos[editar]
- "De l'eau à la terre, une histoire d'évolution", Le Temps d'un Bivouac, France Inter, 21 de agosto de 2019
- Cyril Langlois (16 mai 2012). «Le développement de la végétation continentale de l'Ordovicien au Dévonien et ses conséquences géologiques». planet-terre.ens-lyon.fr.
| Control de autoridades |
|
|---|
 Datos: Q2646445
Datos: Q2646445