Silurien

Gothlandien, Silurien s.s.
| Notation chronostratigraphique | S |
|---|---|
| Notation française | s |
| Notation RGF | s |
| Équivalences | Le Silurien s.l. inclut l'Ordovicien |
| Niveau | Période / Système |
| Érathème / Ère - Éonothème / Éon | Paléozoïque Phanérozoïque |
Stratigraphie
| Début | Fin |
|---|---|
(Extinction Ordovicien-Silurien) |
Paléogéographie et climat

| Taux de O2 atmosphérique | env. 14 %vol[1] (70 % de l'actuel) |
|---|---|
| Taux de CO2 atmosphérique | env. 4 500 ppm[2] (16 fois le niveau d'avant la révolution industrielle) |
| Température moyenne | 17 °C[3] (+3 °C par rapport à l'actuel) |
| Niveau moyen des mers | +180 m (par rapport à l'actuel)[4] |
- fin de l'orogenèse calédonienne
- 440 Ma : formation du sol par les algues
Faune et flore

- 440 Ma : sortie de l'eau des algues
- 435 Ma : arachnides
- 430 Ma : gnathostomes (vertébrés à mâchoire)
- 430 Ma : appendices pairs antérieurs des vertébrés
- 428 Ma : myriapodes
- 420 Ma : ostéichtyens (vertébrés à squelette minéralisé)
- 420 Ma : actinoptérygiens (poissons à nageoires rayonnés)
- 420 Ma : sortie de l'eau des plantes vasculaires
Le Silurien, parfois anciennement nommé Gothlandien, est un système géologique qui s'étend de −443,4 à −419,2 Ma. La datation de début et de fin, bien que définie avec précision par les couches stratigraphiques de référence, est connue à seulement quelques millions d'années près. Il est précédé de l'Ordovicien et suivi du Dévonien. La fin de l'Ordovicien est marquée par l'une des cinq grandes extinctions massives de l'histoire des temps géologiques, où près de 60 % des espèces marines ont disparu et peut-être 85 % du total des espèces (végétales et animales) : l'extinction Ordovicien-Silurien.
Le Silurien a été décrit par Roderick Murchison en 1839 (The Silurian System) se basant sur des terrains découverts dans le sud du Pays de Galles. Il a été nommé en 1835 d'après une tribu celtique : les Silures. Cette classification est entrée en conflit avec celle d'Adam Sedgwick du Cambrien ; ce problème a été résolu par Charles Lapworth en ajoutant l'Ordovicien entre le Cambrien et le Silurien pour les couches géologiques contestées. Le nom « Gothlandien » provient des gisements de l'île suédoise de Gotland.
Subdivisions
[modifier | modifier le code]Le Silurien est divisé en quatre séries ou époques, bien que certains auteurs le découpaient autrefois en trois (Silurien supérieur, moyen et inférieur). Les datations des subdivisions ont été revues par la Commission internationale de stratigraphie (ICS) en 2012[5].
| Pridolien | (423,0 ± 2,3 – 419,2 ± 3,2 Ma) |
| Ludlowien | |
| Ludfordien | (425,6 ± 0,9 – 423,0 ± 2,3 Ma) |
| Gorstien | (427,4 ± 0,5 – 425,6 ± 0,9 Ma) |
| Wenlockien | |
| Homérien | (430,5 ± 0,7 – 427,4 ± 0,5 Ma) |
| Sheinwoodien | (433,4 ± 0,8 – 430,5 ± 0,7 Ma) |
| Llandoveryen | |
| Télychien | (438,5 ± 1,1 – 433,4 ± 0,8 Ma) |
| Aéronien | (440,8 ± 1,2 – 438,5 ± 1,1 Ma) |
| Rhuddanien | (443,4 ± 1,5 – 440,8 ± 1,2 Ma) |
Paléogéographie
[modifier | modifier le code]Pendant le Silurien, le Gondwana reste dans les latitudes hautes de l'hémisphère sud mais la calotte glaciaire est moins étendue que pendant l'Ordovicien. Les autres continents se rapprochent pour commencer la formation d'un second supercontinent : la Laurussia, ce qui ferme l'océan Iapetus qui séparait les divers continents de l'hémisphère nord avant leur accrétion. Ceci a donné naissance à l'orogenèse calédonienne dont on retrouve les traces en Scandinavie. Cette grande chaine de montagnes se poursuit vers l'actuel sud-ouest vers le Canada et les États-Unis où elle forme les Appalaches.
Les petits terranes accrétés au Gondwana pendant l'Ordovicien se détachent à nouveau pour former le superterrane hunnique, appelé ainsi car constitué de la majeure partie des territoires dévastés par Attila[6]. Des couches d'hématite — un minerai de fer — ont été importantes pour l'économie en Amérique du Nord.
Faune silurienne
[modifier | modifier le code]La vie est abondante notamment en milieu marin, car le niveau des océans est élevé en Europe et dans le nord-est de l'Amérique. Toutefois des plantes multicellulaires encore simples, à spores, colonisent les terres, accompagnées par des arthropodes. Les poissons se sont diversifiés considérablement et ont développé des écailles mobiles. Une faune diverse de scorpions de mer, certains longs de plusieurs mètres, se trouve en Amérique du Nord. Échinodermes, mollusques et graptolites sont toujours abondants.
Le Silurien en France
[modifier | modifier le code]À la base des schistes verts de l'unité de Génis, dans le Bas Limousin, apparaissent des lentilles d'un calcaire à entroques contenant des conodontes du Silurien supérieur[7].
Un transect géologique au niveau de la région du houtland en Flandre montre un socle de schistes datant du Silurien vers 300–400 m de profondeur[8].
Climat et niveau de la mer
[modifier | modifier le code]On pensait autrefois que la période du Silurien avait bénéficié de températures relativement stables et chaudes, contrairement aux glaciations extrêmes de l'Ordovicien qui l'avaient précédé et à la chaleur extrême du Dévonien qui a suivi ; cependant, on sait maintenant que le climat mondial a subi de nombreuses fluctuations drastiques tout au long du Silurien[9],[10], comme en témoignent de nombreuses excursions majeures des isotopes du carbone et de l'oxygène au cours de cette période géologique[11],[12],[13]. Le niveau de la mer s'est élevé à partir de son minimum hirnantien tout au long de la première moitié du Silurien ; ils ont ensuite diminué pendant le reste de la période, même si des tendances à plus petite échelle se superposent à cette tendance générale ; quinze hauts plateaux (périodes pendant lesquelles le niveau de la mer était au-dessus du bord du plateau continental) peuvent être identifiés, et le niveau de la mer le plus élevé du Silurien était probablement d'environ 140 mètres plus haut que le niveau le plus bas atteint[14].
Au cours de cette période, la Terre est entrée dans une phase de serre chaude, soutenue par des niveaux élevés de CO2 de 4 500 ppm, et des mers chaudes et peu profondes couvraient une grande partie des masses terrestres équatoriales[15]. Au début du Silurien, les glaciers se sont retirés vers le pôle Sud jusqu'à ce qu'ils disparaissent presque au milieu du Silurien[10]. Les couches de coquilles brisées (appelées coquina) fournissent des preuves solides d'un climat dominé par de violentes tempêtes générées alors comme aujourd'hui par les surfaces marines chaudes[16].
Perturbations
[modifier | modifier le code]Le climat et le cycle du carbone semblent plutôt instables au cours du Silurien, qui présentait une fréquence d'excursions isotopiques (indicatrice de fluctuations climatiques) plus élevée que toute autre période[14]. L’événement Ireviken, l’événement Mulde et l’événement Lau représentent chacun des excursions isotopiques consécutives à une extinction massive mineure[17] et associées à un changement rapide du niveau de la mer. Chacun laisse une signature similaire dans les archives géologiques, à la fois géochimiques et biologiques ; les organismes pélagiques (nageant librement) ont été particulièrement durement touchés, tout comme les brachiopodes, les coraux et les trilobites, et les extinctions se produisent rarement dans une série rapide d'éclats rapides[14],[13]. Les fluctuations climatiques s'expliquent mieux par une séquence de glaciations, mais le manque de tillites du Silurien moyen à supérieur rend cette explication problématique[18].
Flore et faune
[modifier | modifier le code]La période silurienne a été considérée par certains paléontologues comme un intervalle de récupération prolongé après l'extinction massive de l'Ordovicien supérieur (LOME, pour l'anglais Late Ordovician Mass Extinction), qui a interrompu l'augmentation en cascade de la biodiversité qui s'était continuellement poursuivie tout au long du Cambrien et de la majeure partie de l'Ordovicien[19].
Le Silurien a été la première période à voir des mégafossiles d'un vaste biote terrestre sous la forme de forêts miniatures ressemblant à de la mousse le long des lacs et des ruisseaux et des réseaux de grands nématophytes mycorhiziens, annonçant le début de la révolution terrestre silurienne-dévonienne[20],[21],[22]. Cependant, la faune terrestre n'a pas eu d'impact majeur sur la Terre jusqu'à ce qu'elle se diversifie au Dévonien[14].
Les premières traces fossiles de plantes vasculaires, c'est-à-dire de plantes terrestres dont les tissus transportent l'eau et la nourriture, sont apparues dans la seconde moitié de la période silurienne[23]. Les premiers représentants connus de ce groupe sont Cooksonia. La plupart des sédiments contenant Cooksonia sont de nature marine. Les habitats préférés se trouvaient probablement le long des rivières et des ruisseaux. Baragwanathia semble être presque aussi vieux, datant du début de Ludlow (420 millions d'années) et a des tiges ramifiées et des feuilles en forme d'aiguilles de 10 à 20 centimètres. La plante présente un haut degré de développement par rapport à l'âge de ses restes fossiles. Des fossiles de cette plante ont été enregistrés en Australie[24],[25] au Canada[26] et en Chine[27]. Eohostimella heathana est une « plante » précoce, probablement terrestre, connue à partir de fossiles de compression[28] de l'âge du Silurien inférieur (Llandovery)[29].[43] La chimie de ses fossiles est similaire à celle des plantes vasculaires fossilisées, plutôt qu'à celle des algues[28].
Des fossiles considérés comme des animaux terrestres sont également connus du Silurien. Le plus ancien signalement définitif de mille-pattes jamais connu est Kampecaris obanensis et Archidesmus sp. du Silurien supérieur (il y a 425 millions d'années) de Kerrera[30]. Il existe également d'autres mille-pattes, mille-pattes et arachnoïdes trigonotarbidés connus à Ludlow (il y a 420 millions d'années)[30],[31],[32]. Les invertébrés prédateurs indiqueraient qu’il existe des réseaux trophiques simples comprenant des proies non prédatrices. En extrapolant à partir du biote du Dévonien inférieur, Andrew Jeram et al. en 1990[33] a suggéré un réseau trophique basé sur des détritivores et des brouteurs de micro-organismes encore inconnus[34]. Les mille-pattes de la formation de Cowie tels que Cowiedesmus et Pneumodesmus étaient considérés comme les mille-pattes les plus anciens du Silurien moyen il y a 428 millions d'années[35],[36]. Bien que l'âge de cette formation soit plus tard réinterprété comme étant celui du Dévonien inférieur par certains chercheurs[37], une étude plus ultérieure en 2023 a reconfirmé son âge comme étant la fin de l'époque Wenlock du Silurien moyen[38]. Quoi qu'il en soit, Pneumodesmus est toujours un fossile important en tant que preuve définitive la plus ancienne de stigmates permettant de respirer dans l'air.
Apparurent les premiers poissons osseux, les Osteichthyes, représentés par les Acanthodiens couverts d'écailles osseuses. Les poissons ont atteint une diversité considérable et ont développé des mâchoires mobiles, adaptées des supports des deux ou trois arcs branchiaux antérieurs. Une faune diversifiée d'euryptérides (scorpions marins), certains mesurant plusieurs mètres de long, rôdait dans les mers et les lacs peu profonds du Silurien d'Amérique du Nord ; bon nombre de leurs fossiles ont été découverts dans l’État de New York. Les sangsues ont également fait leur apparition au cours de la période silurienne. Les brachiopodes étaient abondants et diversifiés, la composition taxonomique, l'écologie et la biodiversité des brachiopodes du Silurien reflétant celles de l'Ordovicien[39]. Les brachiopodes qui ont survécu à LOME ont développé de nouvelles adaptations au stress environnemental[40] et ont eu tendance à être endémiques à une seule paléoplaque au lendemain de l'extinction massive, mais ont ensuite élargi leur aire de répartition[41]. Les brachiopodes les plus abondants étaient les atrypides et les pentamrides[42] ; les atrypides ont été les premiers à se rétablir et à se rediversifier dans le Rhuddanien après LOME[43], tandis que la récupération des pentamérides a été retardée jusqu'à l'Aéronien[42]. Les bryozoaires présentaient des degrés significatifs d'endémisme sur un plateau particulier[44]. Ils ont également développé des relations symbiotiques avec les cnidaires[45] et les stromatolites[46]. De nombreux fossiles de bivalves ont également été trouvés dans les gisements du Silurien[47] et les premiers bivalves profonds sont connus de cette période[48]. Les chitons ont connu un pic de diversité au milieu du Silurien[49]. Les hédérelloïdes ont connu un succès significatif au Silurien, avec quelques symbioses développantes avec le corail rugueux colonial Entelophyllum[50]. Le Silurien a été l'apogée des tentaculitoïdes[51], qui ont connu un rayonnement évolutif concentré principalement en Baltoscandie[52], ainsi qu'une expansion de leur aire de répartition géographique dans le Llandovery et le Wenlock[53]. Les trilobites ont commencé à se rétablir au Rhuddanien[54] et ont continué à connaître du succès au Silurien comme ils l'avaient fait à l'Ordovicien malgré la réduction de la diversité des clades à la suite de LOME[55]. Le Silurien inférieur a été une période chaotique de renouvellement pour les crinoïdes alors qu'ils se rediverversaient après LOME[56]. Les membres de Flexibilia, qui ont été peu impactés par LOME, ont pris une importance écologique croissante dans les mers du Silurien[57]. Les camerates monobathridés, comme les flexibles, se sont diversifiés dans le Llandovery, tandis que les cyathocrinidés et les dendrocrinidés se sont diversifiés plus tard au Silurien[58]. Les lobolithes scyphocrinoïdes sont soudainement apparus dans le Silurien terminal, peu avant la limite Silurien-Dévonien, et ont disparu aussi brusquement qu'ils sont apparus très peu de temps après leur première apparition[59]. Les symbiotes endobiotiques étaient courants dans les coraux et les stromatoporoïdes[60],[61]. Les coraux rugueux en particulier étaient colonisés et incrustés par une gamme diversifiée d'épibiontes[62], y compris certains hédérelloïdes comme mentionné ci-dessus[50]. Les scléractiniens photosymbiotiques ont fait leur première apparition au Silurien moyen[63]. L'abondance des récifs était inégale ; parfois, les fossiles sont fréquents, mais à d'autres endroits, ils sont pratiquement absents des archives rocheuses[14].
-
 Cooksonia, la première plante vasculaire, Silurien moyen
Cooksonia, la première plante vasculaire, Silurien moyen -
 Fossiles des fonds marins du Silurien collectés dans la réserve naturelle de Wren's Nest, Dudley UK
Fossiles des fonds marins du Silurien collectés dans la réserve naturelle de Wren's Nest, Dudley UK -
 Fragments de crinoïdes dans un calcaire du Silurien (Pridoli) (Saaremaa, Estonie)
Fragments de crinoïdes dans un calcaire du Silurien (Pridoli) (Saaremaa, Estonie) -
 Fossiles des fonds marins du Silurien collectés dans la réserve naturelle de Wren's Nest, Dudley UK
Fossiles des fonds marins du Silurien collectés dans la réserve naturelle de Wren's Nest, Dudley UK -
 Eurypterus, un euryptéride commun du Silurien supérieur
Eurypterus, un euryptéride commun du Silurien supérieur -
 Pterygotus était un eurypteridé géant qui avait une distribution presque cosmopolite (la reconstruction présentée ici est l'espèce dévonienne P. anglicus)
Pterygotus était un eurypteridé géant qui avait une distribution presque cosmopolite (la reconstruction présentée ici est l'espèce dévonienne P. anglicus) -
 Les trilobites étaient encore divers et communs au Silurien. Les fossiles de Calymene celebra sont extrêmement abondants dans certaines parties du centre des États-Unis.
Les trilobites étaient encore divers et communs au Silurien. Les fossiles de Calymene celebra sont extrêmement abondants dans certaines parties du centre des États-Unis. -
 Halysites étaient un corail tabulé, un groupe éteint qui a vécu au Paléozoïque.
Halysites étaient un corail tabulé, un groupe éteint qui a vécu au Paléozoïque. -
 Parioscorpio était un arthropode énigmatique du Silurien du Wisconsin.
Parioscorpio était un arthropode énigmatique du Silurien du Wisconsin. -
 Un spécimen de Dalmanites limulurus Dalmanites limulurus provenant des strates siluriennes de New York
Un spécimen de Dalmanites limulurus Dalmanites limulurus provenant des strates siluriennes de New York -
 Une roche contenant plusieurs brachiopodes pentaméridés géodisés provenant de strates de l'Indiana.
Une roche contenant plusieurs brachiopodes pentaméridés géodisés provenant de strates de l'Indiana. -
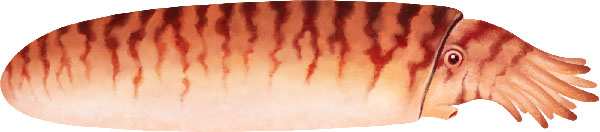 Sphooceras était un céphalopode nautiloïde trouvé dans les strates siluriennes de la République tchèque
Sphooceras était un céphalopode nautiloïde trouvé dans les strates siluriennes de la République tchèque -
 Jamoytius était un vertébré énigmatique qui pourrait être apparenté au poisson Anaspid.
Jamoytius était un vertébré énigmatique qui pourrait être apparenté au poisson Anaspid. -
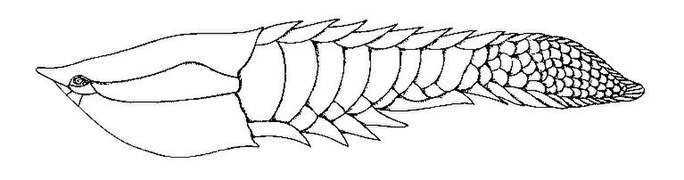 Poraspis, un genre de poisson blindé sans mâchoire du Silurien supérieur du Canada, de la Norvège et des États-Unis.
Poraspis, un genre de poisson blindé sans mâchoire du Silurien supérieur du Canada, de la Norvège et des États-Unis. -
 Tujiaaspis est un agnathan galeaspid du début du Silurien (Telychian) de Chine, montrant l'origine de nageoires appariées
Tujiaaspis est un agnathan galeaspid du début du Silurien (Telychian) de Chine, montrant l'origine de nageoires appariées -
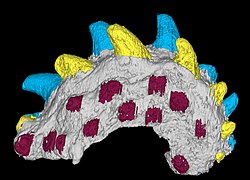
-
 Fanjingshania, requin épineux climatiidé du Silurien inférieur (Aéronien) décrit à partir d'éléments dermosquelettiques désarticulés
Fanjingshania, requin épineux climatiidé du Silurien inférieur (Aéronien) décrit à partir d'éléments dermosquelettiques désarticulés -
 Shenacanthus[65] est un genre de Chondrichtyen à tige mâchoire du Silurien inférieur (Telychien) de Chine.
Shenacanthus[65] est un genre de Chondrichtyen à tige mâchoire du Silurien inférieur (Telychien) de Chine. -
![Xiushanosteus[65] is the oldest known placoderm from the early Silurian (Telychian) of China](//upload.wikimedia.org/wikipedia/commons/thumb/c/c5/Xiushanosteus.jpg/191px-Xiushanosteus.jpg) Xiushanosteus[65] is the oldest known placoderm from the early Silurian (Telychian) of China
Xiushanosteus[65] is the oldest known placoderm from the early Silurian (Telychian) of China -
 Entelognathus primordialis est le plus ancien placoderme connu du début du Silurien (Telychien) de Chine.
Entelognathus primordialis est le plus ancien placoderme connu du début du Silurien (Telychien) de Chine.










_1.jpg)







![Xiushanosteus[65] is the oldest known placoderm from the early Silurian (Telychian) of China](Fichier:Xiushanosteus.jpg)

Voir aussi
[modifier | modifier le code]Articles connexes
[modifier | modifier le code]Liens externes
[modifier | modifier le code]Notes et références
[modifier | modifier le code]Notes
[modifier | modifier le code]- Notices dans des dictionnaires ou encyclopédies généralistes :
Références
[modifier | modifier le code]- (de) teneur en oxygène dans l'atmosphère au Phanérozoïque
- (en) dioxyde de carbone au Phanérozoïque
- (en) température de la Terre
- (fr) variation du niveau des mers au Phanérozoïque
- [PDF] (en) « International chronostratigraphic chart (2012) », sur stratigraphy.org.
- (en) Gérard M. Stampfli, « Tethyan oceans », Geological Society, London, Special Publications, vol. 173, , p. 1-23 (DOI 10.1144/GSL.SP.2000.173.01.01).
- Guillot PL. & Lefebvre J.J., 1975. Découverte de conodontes dans le calcaire à entroques de Génis en Dordogne (série métamorphique du Bas Limousin). C. R. Acad. Sci., 280, pp. 1529-1530
- Géologie en Nord - Pas de Calais sur le Système d’information pour la gestion des eaux souterraines en Nord-Pas de Calais.
- Guanzhou Yan, Oliver Lehnert, Peep Männik, Mikael Calner, Xiaocong Luan, Fanyi Gong, Lixia Li, Xin Wei, Guangxu Wang, Renbin Zhan et Rongchang Wu, « The record of early Silurian climate changes from South China and Baltica based on integrated conodont biostratigraphy and isotope chemostratigraphy », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 606, , p. 111245 (DOI 10.1016/j.palaeo.2022.111245, Bibcode 2022PPP...60611245Y, S2CID 252504361, lire en ligne, consulté le )
- G. Gambacorta, E. Menichetti, E. Trincianti et S. Torricelli, « The Silurian climatic transition recorded in the epicontinental Baltica Sea », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 517, , p. 16–29 (DOI 10.1016/j.palaeo.2018.12.016, Bibcode 2019PPP...517...16G, S2CID 135118794)
- Set A. Young, Emily Benayoun, Nevin P. Kozik, Olle Hints, Tõnu Martma, Stig M. Bergström et Jeremy D. Owens, « Marine redox variability from Baltica during extinction events in the latest Ordovician–early Silurian », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 554, , p. 109792 (DOI 10.1016/j.palaeo.2020.109792
 , Bibcode 2020PPP...55409792Y, S2CID 218930512)
, Bibcode 2020PPP...55409792Y, S2CID 218930512) - Adam D. Sproson, « Pacing of the latest Ordovician and Silurian carbon cycle by a ~4.5 Myr orbital cycle », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 540, , p. 109543 (DOI 10.1016/j.palaeo.2019.109543, Bibcode 2020PPP...54009543S, S2CID 213445668, lire en ligne, consulté le )
- Julie A. Trotter, Ian S. Williams, Christopher R. Barnes, Peep Männik et Andrew Simpson, « New conodont δ18O records of Silurian climate change: Implications for environmental and biological events », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 443, , p. 34–48 (DOI 10.1016/j.palaeo.2015.11.011, Bibcode 2016PPP...443...34T)
- Axel Munnecke, Mikael Calner, David A.T. Harper et Thomas Servais, « Ordovician and Silurian sea–water chemistry, sea level, and climate: A synopsis », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 296, nos 3–4, , p. 389–413 (DOI 10.1016/j.palaeo.2010.08.001, Bibcode 2010PPP...296..389M)
- William G. Chaloner, « The role of carbon dioxide in plant evolution », Evolution on Planet Earth, , p. 65–83 (ISBN 9780125986557, DOI 10.1016/B978-012598655-7/50032-X)
- T. Nealon et D. Michael Williams, « Storm-influenced shelf deposits from the silurian of Western Ireland: A reinterpretation of deep basin sediments », Geological Journal, vol. 23, no 4, , p. 311–320 (DOI 10.1002/gj.3350230403)
- C. Samtleben, A. Munnecke et T. Bickert, « Development of facies and C/O-isotopes in transects through the Ludlow of Gotland: Evidence for global and local influences on a shallow-marine environment », Facies, vol. 43, , p. 1–38 (DOI 10.1007/BF02536983, S2CID 130640332)
- Mikael Calner, Mass Extinction, , 21–57 p. (ISBN 978-3-540-75915-7, DOI 10.1007/978-3-540-75916-4_4), « Silurian global events – at the tipping point of climate change »
- Christian M. Ø. Rasmussen, Björn Kröger, Morten L. Nielsen et Jorge Colmenar, « Cascading trend of Early Paleozoic marine radiations paused by Late Ordovician extinctions », Proceedings of the National Academy of Sciences of the United States of America, vol. 116, no 15, , p. 7207–7213 (PMID 30910963, PMCID 6462056, DOI 10.1073/pnas.1821123116 , Bibcode 2019PNAS..116.7207R)
- Elliot Capel, Christopher J. Cleal, Jinzhuang Xue, Claude Monnet, Thomas Servais et Borja Cascales-Miñana, « The Silurian–Devonian terrestrial revolution: Diversity patterns and sampling bias of the vascular plant macrofossil record », Earth-Science Reviews, vol. 231, , p. 104085 (DOI 10.1016/j.earscirev.2022.104085 , Bibcode 2022ESRv..23104085C, hdl 20.500.12210/76731 , S2CID 249616013)
- Jinzhuang Xue, Pu Huang, Deming Wang, Conghui Xiong, Le Liu et James F. Basinger, « Silurian-Devonian terrestrial revolution in South China: Taxonomy, diversity, and character evolution of vascular plants in a paleogeographically isolated, low-latitude region », Earth-Science Reviews, vol. 180, , p. 92–125 (DOI 10.1016/j.earscirev.2018.03.004, Bibcode 2018ESRv..180...92X, lire en ligne, consulté le )
- Gregory J. Retallack, « Ordovician-Devonian lichen canopies before evolution of woody trees », Gondwana Research, vol. 106, , p. 211–223 (DOI 10.1016/j.gr.2022.01.010, Bibcode 2022GondR.106..211R, S2CID 246320087, lire en ligne, consulté le )
- Don Rittner, Encyclopedia of Biology, Infobase Publishing, (ISBN 9781438109992, lire en ligne), p. 338
- R. B. Rickards, « The age of the earliest club mosses: the Silurian Baragwanathia flora in Victoria, Australia », Geological Magazine, vol. 137, no 2, , p. 207–209 (DOI 10.1017/S0016756800003800, Bibcode 2000GeoM..137..207R, S2CID 131287538, lire en ligne, consulté le )
- W.H. Lang et I.C. Cookson, « On a flora, including vascular land plants, associated with Monograptus, in rocks of Silurian age, from Victoria, Australia », Philosophical Transactions of the Royal Society of London B, vol. 224, no 517, , p. 421–449 (DOI 10.1098/rstb.1935.0004 , Bibcode 1935RSPTB.224..421L)
- F.M. Hueber, « A new species of Baragwanathia from the Sextant Formation (Emsian) Northern Ontario, Canada », Botanical Journal of the Linnean Society, vol. 86, nos 1–2, , p. 57–79 (DOI 10.1111/j.1095-8339.1983.tb00717.x)
- Lily Bora, Principles of Paleobotany, Mittal Publications, , 36–37 p.
- Niklas, Karl J., « Chemical Examinations of Some Non-Vascular Paleozoic Plants », Brittonia, vol. 28, no 1, , p. 113–137 (DOI 10.2307/2805564, JSTOR 2805564, S2CID 21794174)
- D. Edwards et C. Wellman, Plants Invade the Land : Evolutionary and Environmental Perspectives, New York: Columbia University Press, , 3–28 p. (ISBN 978-0-231-11161-4), p. 4
- (en) M. E. Brookfield, E. J. Catlos et S. E. Suarez, « Myriapod divergence times differ between molecular clock and fossil evidence: U/Pb zircon ages of the earliest fossil millipede-bearing sediments and their significance », Historical Biology, vol. 33, no 10, , p. 2014–2018 (ISSN 0891-2963, DOI 10.1080/08912963.2020.1762593, S2CID 238220137, lire en ligne)
- Jason A. Dunlop, « A trigonotarbid arachnid from the Upper Silurian of Shropshire », Palaeontology, vol. 39, , p. 605–614 (ISSN 0031-0239, lire en ligne)
- William A. Shear, Andrew J. Jeram et Paul Selden, Centiped legs (Arthropoda, Chilopoda, Scutigeromorpha) from the Silurian and Devonian of Britain and the Devonian of North America, New York, NY, American Museum of Natural History, (lire en ligne)
- Andrew J. Jeram, Paul A. Selden et Dianne Edwards, « Land Animals in the Silurian: Arachnids and Myriapods from Shropshire, England », Science, vol. 250, no 4981, , p. 658–61 (PMID 17810866, DOI 10.1126/science.250.4981.658, Bibcode 1990Sci...250..658J)
- William A DiMichele et Robert W Hook, Terrestrial Ecosystems Through Time: Evolutionary Paleoecology of Terrestrial Plants and Animals, University of Chicago Press, , 207–10 p. (ISBN 978-0-226-04155-1), « The Silurian »
- Paul Selden & Helen Read, « The oldest land animals: Silurian millipedes from Scotland », Bulletin of the British Myriapod & Isopod Group, vol. 23, , p. 36–37 (lire en ligne)
- (en) Heather M. Wilson et Lyall I. Anderson, « Morphology and taxonomy of Paleozoic millipedes (Diplopoda: Chilognatha: Archipolypoda) from Scotland », Journal of Paleontology, vol. 78, no 1, , p. 169–184 (ISSN 0022-3360, DOI 10.1666/0022-3360(2004)078<0169:MATOPM>2.0.CO;2, S2CID 131201588, lire en ligne)
- Stephanie E. Suarez, Michael E. Brookfield, Elizabeth J. Catlos et Daniel F. Stöckli, « A U-Pb zircon age constraint on the oldest-recorded air-breathing land animal », PLOS ONE, vol. 12, no 6, , e0179262 (ISSN 1932-6203, PMID 28658320, PMCID 5489152, DOI 10.1371/journal.pone.0179262 )
- C.H. Wellman, G. Lopes, Z. McKellar et A. Hartley, « Age of the basal 'Lower Old Red Sandstone' Stonehaven Group of Scotland: The oldest reported air-breathing land animal is Silurian (late Wenlock) in age », The Geological Society of London, vol. 181, (ISSN 0016-7649, DOI 10.1144/jgs2023-138 , hdl 2164/22754 )
- Jia-yu Rong et Shu-zhong Shen, « Comparative analysis of the end-Permian and end-Ordovician brachiopod mass extinctions and survivals in South China », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 188, nos 1–2, , p. 25–38 (DOI 10.1016/S0031-0182(02)00507-2, Bibcode 2002PPP...188...25R, lire en ligne, consulté le )
- Jiayu Rong et Renbin Zhan, « Chief sources of brachiopod recovery from the end Ordovician mass extinction with special references to progenitors », Science in China Series D: Earth Sciences, vol. 42, no 5, , p. 553–560 (DOI 10.1007/BF02875250, Bibcode 1999ScChD..42..553R, S2CID 129323463, lire en ligne, consulté le )
- Bing Huang, Jiayu Rong et L. Robin M. Cocks, « Global palaeobiogeographical patterns in brachiopods from survival to recovery after the end-Ordovician mass extinction », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 317-318, , p. 196–205 (DOI 10.1016/j.palaeo.2012.01.009, Bibcode 2012PPP...317..196H, lire en ligne, consulté le )
- Bing Huang, Jisuo Jin et Jia-Yu Rong, « Post-extinction diversification patterns of brachiopods in the early–middle Llandovery, Silurian », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 493, , p. 11–19 (DOI 10.1016/j.palaeo.2017.12.025, Bibcode 2018PPP...493...11H, lire en ligne, consulté le )
- Rong Jia-Yu et David A. T. Harper, « Brachiopod survival and recovery from the latest Ordovician mass extinctions in South China », Geological Journal, vol. 34, no 4, , p. 321–348 (DOI 10.1002/(SICI)1099-1034(199911/12)34:4<321::AID-GJ809>3.0.CO;2-I, lire en ligne, consulté le )
- Robert L. Anstey, Joseph F. Pachut et Michael E. Tuckey, « Patterns of bryozoan endemism through the Ordovician-Silurian transition », Paleobiology, vol. 29, no 3, , p. 305–328 (DOI 10.1666/0094-8373(2003)0292.0.CO;2, lire en ligne, consulté le )
- Mikołaj K. Zapalski, Olev Vinn, Ursula Toom, Andrej Ernst et Mark A. Wilson, « Bryozoan–cnidarian mutualism triggered a new strategy for greater resource exploitation as early as the Late Silurian », Scientific Reports, vol. 12, no 1, , p. 15556 (PMID 36114227, PMCID 9481587, DOI 10.1038/s41598-022-19955-2, Bibcode 2022NatSR..1215556Z)
- Anna Lene Claussen, Axel Munnecke et Andrej Ernst, « Bryozoan-rich stromatolites (bryostromatolites) from the Silurian of Gotland and their relation to climate-related perturbations of the global carbon cycle », Sedimentology, vol. 69, no 1, , p. 162–198 (DOI 10.1111/sed.12863 , S2CID 233820331)
- Louis Liljedahl, « Ecological aspects of a silicified bivalve fauna from the Silurian of Gotland », Lethaia, vol. 18, no 1, , p. 53–66 (DOI 10.1111/j.1502-3931.1985.tb00684.x, lire en ligne, consulté le )
- Anna Lene Claussen, Axel Munnecke, Mark A. Wilson et Irina Oswald, « The oldest deep-boring bivalves? Evidence from the Silurian of Gotland (Sweden) », Facies, vol. 65, no 3, (DOI 10.1007/s10347-019-0570-7, S2CID 149500698, lire en ligne, consulté le )
- Lesley Cherns, « Early Palaeozoic diversification of chitons (Polyplacophora, Mollusca) based on new data from the Silurian of Gotland, Sweden », Lethaia, vol. 37, no 4, , p. 445–456 (DOI 10.1080/00241160410002180, lire en ligne, consulté le )
- Olev Vinn, Mark A. Wilson, Anna Madison, Elizaveta Kazantseva et Ursula Toom, « First Symbiotic Association Between Hederelloids and Rugose Corals (Latest Silurian of Saaremaa, Estonia) », PALAIOS, vol. 37, no 7, , p. 368–373 (DOI 10.2110/palo.2022.005, Bibcode 2022Palai..37..368V, S2CID 251122025, lire en ligne, consulté le )
- Olev Vinn, « Adaptive strategies in the evolution of encrusting tentaculitoid tubeworms », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 292, nos 1–2, , p. 211–221 (DOI 10.1016/j.palaeo.2010.03.046, Bibcode 2010PPP...292..211V, lire en ligne, consulté le )
- Jacalyn M. Wittmer et Arnold I. Miller, « Dissecting the global diversity trajectory of an enigmatic group: The paleogeographic history of tentaculitoids », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 312, nos 1–2, , p. 54–65 (DOI 10.1016/j.palaeo.2011.09.009, Bibcode 2011PPP...312...54W, lire en ligne, consulté le )
- Fan Wei, Yiming Gong et Hao Yang, « Biogeography, ecology and extinction of Silurian and Devonian tentaculitoids », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 358-360, , p. 40–50 (DOI 10.1016/j.palaeo.2012.07.019, Bibcode 2012PPP...358...40W, lire en ligne, consulté le )
- Xin Wei et Ren-Bin Zhan, « A late Rhuddanian (early Llandovery, Silurian) trilobite association from South China and its implications », Palaeoworld, vol. 27, no 1, , p. 42–52 (DOI 10.1016/j.palwor.2017.05.008, lire en ligne, consulté le )
- Jonathan M. Adrain, Stephen R. Westrop, Brian D. E. Chatterton et Lars Ramsköld, « Silurian trilobite alpha diversity and the end-Ordovician mass extinction », Paleobiology, vol. 26, no 4, , p. 625–646 (DOI 10.1666/0094-8373(2000)0262.0.CO;2, lire en ligne, consulté le )
- Shanan E. Peters et William I. Ausich, « A sampling-adjusted macroevolutionary history for Ordovician-Early Silurian crinoids », Paleobiology, vol. 34, no 1, , p. 104–116 (DOI 10.1666/07035.1, S2CID 67838459, lire en ligne, consulté le )
- David F. Wright et Ursula Toom, « New crinoids from the Baltic region (Estonia): fossil tip-dating phylogenetics constrains the origin and Ordovician–Silurian diversification of the Flexibilia (Echinodermata) », Palaeontology, vol. 60, no 6, , p. 893–910 (DOI 10.1111/pala.12324 , S2CID 134049045)
- William I. Ausich et Bradley Deline, « Macroevolutionary transition in crinoids following the Late Ordovician extinction event (Ordovician to Early Silurian) », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 361-362, , p. 38–48 (DOI 10.1016/j.palaeo.2012.07.022, Bibcode 2012PPP...361...38A, lire en ligne, consulté le )
- Rui-wen Zong et Yi-ming Gong, « Discovery of scyphocrinoid loboliths in western Junggar, Xinjiang, NW China: Implications for scyphocrinoid paleobiogeography and identification of the Silurian–Devonian boundary », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 557, , p. 109914 (DOI 10.1016/j.palaeo.2020.109914, Bibcode 2020PPP...55709914Z, S2CID 224964119, lire en ligne, consulté le )
- Vinn, O., wilson, M.A. et Mõtus, M.-A., « Symbiotic endobiont biofacies in the Silurian of Baltica », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 404, , p. 24–29 (DOI 10.1016/j.palaeo.2014.03.041, Bibcode 2014PPP...404...24V, lire en ligne, consulté le )
- Vinn, O. et Mõtus, M.-A., « The earliest endosymbiotic mineralized tubeworms from the Silurian of Podolia, Ukraine », Journal of Paleontology, vol. 82, no 2, , p. 409–414 (DOI 10.1666/07-056.1, Bibcode 2008JPal...82..409V, S2CID 131651974, lire en ligne, consulté le )
- Michał Zatoń et Tomasz Wrzołek, « Colonization of rugose corals by diverse epibionts: dominance and syn vivo encrustation in a Middle Devonian (Givetian) soft-bottom habitat of the Holy Cross Mountains, Poland », Palaeogeography, Palaeoclimatology, Palaeoecology, vol. 556, , p. 109899 (DOI 10.1016/j.palaeo.2020.109899, Bibcode 2020PPP...55609899Z, S2CID 224869051, lire en ligne, consulté le )
- (en) Mikołaj K. Zapalski, « Evidence of photosymbiosis in Palaeozoic tabulate corals », Proceedings of the Royal Society B: Biological Sciences, vol. 281, no 1775, , p. 20132663 (ISSN 0962-8452, PMID 24307674, PMCID 3866410, DOI 10.1098/rspb.2013.2663)
- (en) Plamen S. Andreev, Ivan J. Sansom, Qiang Li, Wenjin Zhao, Jianhua Wang, Chun-Chieh Wang, Lijian Peng, Liantao Jia, Tuo Qiao et Min Zhu, « The oldest gnathostome teeth », Nature, vol. 609, no 7929, , p. 964–968 (ISSN 1476-4687, PMID 36171375, DOI 10.1038/s41586-022-05166-2, Bibcode 2022Natur.609..964A, S2CID 252569771, lire en ligne)
- (en) You-an Zhu, Qiang Li, Jing Lu, Yang Chen, Jianhua Wang, Zhikun Gai, Wenjin Zhao, Guangbiao Wei, Yilun Yu, Per E. Ahlberg et Min Zhu, « The oldest complete jawed vertebrates from the early Silurian of China », Nature, vol. 609, no 7929, , p. 954–958 (ISSN 1476-4687, PMID 36171378, DOI 10.1038/s41586-022-05136-8, S2CID 252569910, lire en ligne)

{kind=link}
{kind=link}
{kind=link}
{kind=link}

| Paléozoïque | Mésozoïque | Cénozoïque | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Cambrien | Ordovicien | Silurien | Dévonien | Carbonifère | Permien | Trias | Jurassique | Crétacé | Paléogène | Néogène | Q. |