Lípid


Els lípids són biomolècules insolubles en aigua que presenten solubilitat elevada en dissolvents orgànics, com ara el cloroform.[1][2]
L'etimologia de la paraula lípid prové de la forma prefixada del mot grec lípos, que significa "greix" i de la forma sufixada del mot grec eĩdos, que significa "aparença, figura".[3]
A diferència dels glúcids i de les proteïnes, que tenen grups funcionals característics, els lípids no tenen cap grup funcional propi. En realitat l'única característica comuna que tenen és la solubilitat dins dissolvents orgànics no polars (benzè, èter dietílic…). Aquesta propietat també els diferencia dels glúcids i les proteïnes que són solubles dins dissolvents polars com l'aigua. La raó per la qual són insolubles en dissolvents polars és perquè les seves estructures químiques estan formades per llargues cadenes de carbonis que són apolars.[4]
Els lípids constitueixen un ampli grup de molècules que es presenten a la natura que inclou els greixos, ceres, esterols, vitamines liposolubles (com les vitamines A, D, E, i K), monoglicèrids, diglicèrids, triacilglicerols, fosfolípids, i d'altres. Les principals funcions dels lípids inclouen l'emmagatzematge d'energia, ser components estructurals de la membrana cel·lular, i com important senyalitzador molecular.[5][6]
A grans trets els lípids es poden definir com petites molècules hidròfobes o amfifíliques; la naturalesa amfifílica d'alguns lípids els permet formar estructures com la vesícula, liposomes, o membranes en un ambient aquós. Els lípids biològics s'originen enterament o en part de dos tipus diferents de subunitats bioquímiques: grups cetoacil i isoprè.[5] Fent servir aquest enfocament, els lípids es poden dividir en 8 categories: àcids grassos, glicerolípids, glicerofosfolípids, esfingolípids, sacarolípids, i poliquètids (derivats de la condensació de subunitats de cetoacil); i lípids esteròlics i lípids prenòlids (derivats de la condensació de subunitats d'isoprè).[5]
Malgrat que el terme lípid de vegades es fa servir com a sinònim de greix, els greixos són un subgrup dels lípids anomenats triacilglicerols. Els lípids també comprenen molècules com els àcids grassos i els seus derivats (incloent tri-, di-, monoglicèrids, i fosfolípids), com també altres metabòlits que contenen esterols com el colesterol.[7] Encara que els humans i altres mamífers fan servir diverses vies metabòliques per degradar i sintetitzar els lípids, alguns lípids essencials no es poden fer d'aquesta manera i s'han d'obtenir de la dieta.
Funcions biològiques a l'organisme
[modifica]
Funció de reserva energètica
[modifica]Els triacilglicerols, emmagatzemats en el teixit adipós, són la principal forma d'emmagatzematge d'energia tant en animals com en plantes. Els triacilglicerols s'acumulen principalment en el teixit adipós de la pell de molts animals i les llavors d'alguns vegetals.
L'adipòcit o cèl·lula de greix, està dissenyat per la síntesi i degradació contínua dels triacilglicerols en els animals, amb la degradació controlada principalment per l'activació de l'enim lipasa.[8] Els lípids, a causa dels àcids grassos que contenen, constitueixen una de les formes més riques en energia de l'alimentació, i llur oxidació a les cèl·lules és completa.
L'oxidació completa dels àcids grassos proporciona un alt contingut calòric, unes 9 kcal/g, comparades amb les 4 kcal/g per la degradació dels glúcids i les proteïnes. Els ocells migradors fan servir l'energia emmagatzemada dels triacilglicerols en els seus vols.[9]
La deposició patològica de lípids a les parets de les artèries intervé en la producció d'un ateroma.
Funció estructural
[modifica]
Certs lípids (fosfolípids i colesterol), són components de la membrana cel·lular de la que formen la bicapa lipídica. Aquesta bicapa és el principal element estructural i també el responsable de la permeabilitat selectiva de la membrana. Les ceres són molt difoses en els insectes, en certs cetacis i en les plantes, en els quals tenen funcions de protecció. En els mamífers, els teixits amb una funció complexa, com el teixit cerebral, el nerviós, l'hepàtic o el renal, tenen el més alt contingut en fosfolípids.
Les cèl·lules dels eucariotes estan compartimentades dins orgànuls units a la membrana que porten a terme diferents funcions biològiques. Els glicerofosfolípids són el principal component estructural de les membranes biològiques, com la membrana plasmàtica cel·lular i les membranes intracel·lulars dels orgànuls; en les cèl·lules animals la membrana plasmàtica separa físicament els components intracel·lulars de l'ambient extracel·lular. Els glicerofosfolípids són molècules amfipàtiques que contenen un nucli de glicerol enllaçat a dues "cues" de dos àcids grassos derivats d'enllaços èster i un "cap" per un enllaç èster fosfat. Mentre que els glicerofosfolípids són el major component de les membranes biològiques, altres components lípids no glicèrids com l'esfingomielina i esterols (principalment colesterol en les membranes cel·lulars dels animals) també es troben en les membranes biològiques.[10] En les plantes i les algues, el galactosildiacilglicerols,[11] sulfoquinovosildiacilglicerol, distromitosidiacilgliercerol[12] el qual no té un grup fosfat, són components importants de les membranes dels cloroplastsi i els orgànuls relacionats i són els lípids més abundants en els teixits fotosintètics, incloent aquells de les plantes superiors, algues i certs bacteris.
S'ha comprovat que les bicapes mostren alts nivells de birefringència, les quals es poden usar per sondejar el grau d'ordre (o disrupció) dins la bicapa usant tècniques com la interferometria a doble polarització i el dicroisme circular.
Una membrana biològica és una forma de bicapa lipídica. La formació de bicapes lipídiques és un procés preferit energèticament quan els glicerofosfolípids estan en ambient aquós.[13] Això es coneix sota el nom d'efecte hidròfob. En un sistema aquós, els caps polars dels lípids s'alineen en direcció al moviment aquós polar, mentre que les cues hidrofòbiques minimitzen el seu contacte amb l'aigua i tendeixen a agrupar-se juntes, formant una vesícula; depenent de la concentració del lípid, aquesta interacció biofísica pot resultar en la formació de micel·les, liposomes, o bicapes lipídiques. Altres agregacions també s'observen i formen part del polimorfisme del comportament del lípids amfífils. El comportament de fase és una àrea d'estudi dins la biofísica i de recerca.[14][15] Les micel·les i les bicapes es formen en un medi polar per l'efecte hidrofòbic.[16] Quan es dissol una substància en un ambient lipofílic o amfifílic, les molècules polars (és a dir aigua en dissolució aquosa) esdevenen més ordenades al voltant de la substància lipofílica, ja que les molècules polars no poden formar enllaços d'hidrogen en les zones lipofíliques de l'amfífil. Per tant en ambient aquós, les molècules d'aigua formen un una gàbia ordenada de clatrat alvoltant de la molècula lipofílica dissolta.[17]
Funció catalítica i reguladora
[modifica]Algunes substàncies que intervenen en nombrosos processos fisiològics són lípids o derivats, com vitamines liposolubles, hormones esteroidals i les prostaglandines.
Funció de transport
[modifica]Els transport de lípids des de l'intestí fins al lloc de destinació és realitzat pels àcids biliars i pels proteolípids.
Senyalització
[modifica]Recentment s'ha vist que la senyalització lipídica és una part vital de la senyalització cel·lular.[18] La senyalització lipídica pot passar via l'activació del receptor de la G-proteïna aparellada o receptors nuclears s'han identificat membres de diferents categories de lípids com a molècules de senyalització i missatgers cel·lulars.[19] Aquests inclouen l'esfingosina-1-fosfat, un esfingolípid derivat de la ceramida que és una potent molècula missatgera implicada en la regulació de la mobilitació del calci,[20] el creixement cel·lular i l'apoptosi;[21] el diacilglicerol (DAG) i els fosfats de fosfatidilisol (PIPs), implicats en l'activació de la proteïna-cinasa C mediada pel calci;[22] les prostaglandines, les quals són un tipus d'àcids grassos implicades en la inflamació i immunitat;[23] les hormones esteroïdes com l'estrogen, testosterona i el cortisol,les quals modulen moltes funcions com la reproducció, el metabolisme i pressió de la sang; i els oxisterols com 25-hidroxi-colesterol que són agonistes de receptor del fetge X.[24]
Altres funcions
[modifica]Les vitamines "solubles en greix" (liposolubles) (A, D, E i K) – les quals són lípids basats en l'isoprè – són nutrients essencials emmagatzemats en el fetge i els teixits adiposos, amb diverses funcions. Les acil-carnitines estan implicades en el transport i el metabolisme dels àcids grassos dins i fora dels mitocondris, on experimenten la beta oxidació.[25] Els poliprenols i els seus derivats fosforilats també tenen papers importants, en el cas dels transport d'oligosacàrids a través de les membranes. Els sucres de fosfat de poliprenol i els sucres de difosfat de poliprenol tenen una funció en les reaccions de glucosilació extracitoplasmàtica, enbiosíntesi de polisacàrids estracel·lular (per exemple la polimerització del peptidolican en bacteris), i en la glucosilació de la proteïna-N en eucariotes.[26][27] Les cardiolipines són una subclasse de glicerofosfolípids que contenen 4 cadenes acil i tres grups glicerol que són particularment abundants en la membrana interior dels mitocondris.[28][29][30] Es creu que activen enzims involucrats en la fosforilació oxidativa.[31] Els lípids també formen la base de les hormones esteroides.[32]
Classificació dels lípids
[modifica]
Els lípids es poden dividir en diversos grups:
Segons la seva composició química
[modifica]- Hom anomena lípids simples els que només contenen carboni, oxigen i hidrogen, i comprenen els triacilglicerols, anomenats greixos o olis segons que siguin sòlids o líquids, les ceres i els èsters dels esterols; dins aquest darrer grup, un dels esterols més coneguts és el colesterol.
- Els lípids complexos contenen, a més, altres elements i són anomenats, en general, lipoides; comprenen els fosfolípids, els esfingolípids i els glicolípids.[33]
Segons la seva capacitat natural de saponificació
[modifica]Una altra classificació es basa en la reacció de saponificació:
- Hom anomena lípids saponificables als lípids que poden reaccionar amb hidròxids alcalins per a formar sabons. Tots contenen àcids grassos. Dins aquest grup hi ha els àcids grassos, els triacilglicerols, els cèrids, els fosfolípids i els icosanoides.
- Els lípids no saponificables són aquells que no reaccionen amb hidròxids alcalins per a donar sabons. Són lípids que no contenen àcids grassos. N'hi ha de dos tipus: Esteroides i terpenoides[4]
Lípids saponificables
[modifica]

Els lípids saponificables són aquells que contenen àcids grassos o derivats. Es diuen així perquè el grup carboxílic de l'àcid gras (o l'èster dels derivats) pot reaccionar amb un hidròxid (NaOH, KOH, etc.) per formar la sal alcalina de l'àcid i alcohol. Històricament, el mot "saponificació" fa referència a la reacció dels greixos amb la potassa càustica o hidròxid de potassi, la qual conduïa a la formació de sabons. Val a dir que, en l'actualitat, més d'un 80% dels sabons usuals són preparats encara per variants d'aquest mètode.
En aquest grup de lípids es troben els àcids grassos, els triacilglicerols, les ceres i els fosfolípids.
Àcids grassos
[modifica]
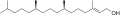
Un àcid gras és un tipus de molècula orgànica formada per una llarga cadena hidrocarbonada en línia, la gran majoria són de nombre parell d'àtoms de carboni i d'entre ells; els més comuns els que tenen 16 i 18,[34] en l'extrem de la qual hi ha un grup carboxil (-COOH). Cada àtom de carboni s'uneix al següent i al precedent per mitjà d'un enllaç covalent senzill.
La fórmula bàsica d'una molècula d'àcid gras saturat és CH3–(CH2)n–COOH. A l'àtom del seu extrem li queden lliures tres enllaços que són ocupats per àtoms d'hidrogen (H3C-). Els altres àtoms tenen lliures dos enllaços, que són ocupats igualment per àtoms d'hidrogen (...-CH2-CH2-CH2-...).
Els àcid grassos que a la cadena de carbonis només hi ha enllaços C-C simples se'ls anomena àcids grassos saturats.


| Àcids grassos saturats més importants | |
| Àcid | Fórmula |
|---|---|
| Àcid làuric | CH3-(CH2)10-COOH |
| Àcid mirístic | CH3-(CH2)12-COOH |
| Àcid palmític | CH3-(CH2)14-COOH |
| Àcid esteàric | CH3-(CH2)16-COOH |
| Àcid araquídic | CH3-(CH2)18-COOH |
| Àcid lignocèric | CH3-(CH2)22-COOH |
Si presenten dobles enllaços C=C se'ls anomena àcid grassos insaturats, i d'aquests n'hi ha de cis i de trans, segons la isomeria del doble enllaç. Els insaturats també poden ser monoinsaturats si només tenen un enllaç doble, com l'àcid oleic, o poliinsaturats si tenen més d'un enllaç doble, com els àcids linoleic i el linolènic. Aquests dos darrers són àcids grassos essencials, és a dir, que el nostre organisme no pot sintetitzar-los i necessita incorporar-los a la dieta a partir d'olis vegetals. En general els àcids grassos saturats es troben en els greixos animals, mentre que els insaturats es troben majoritàriament als olis vegetals.
Àcids grassos omega 3
[modifica]


Els àcids grassos omega 3 constitueixen una família d'àcids grassos que es caracteritzen per tenir el seu primer doble enllaç (insaturació) a la posició tercera començant pel darrer àtom de carboni. Aquest tret estructural distintiu els atorga unes propietats físiques especials i els converteix en precursors insubstituïbles de molècules que els éssers vius utilitzen per a regular determinades funcions circulatòries.
La principal font alimentària d'aquests àcids grassos (i l'única que en conté quantitats nutricionalment significatives) és el peix blau; més concretament el seu greix i el seu oli. Actualment però, aquests àcids grassos han estat incorporats de manera artificial en altres aliments (com ara ous i làctics), oferint fonts alimentàries complementàries al peix blau per a la seva ingesta.
Glicerolípids
[modifica]Els glicerolípids estan compostos principalment de glicerols mono-, di-, i tri-substituïts,[35] el glicerolípid més ben conegut són els trièsters de l'àcid gras glicerol anomenats triacilglicerols. La paraula «triacilglicerol» de vegades es fa servir com a sinònim de triacilglicerol, però això no resulta correcte respecte a aquests compostos, ja que no contenen cap grup hidroxil. En aquests compostos els tres grups hidroxil del glicerol estan esterificats, típicament per diferents àcids grassos. Com que la seva funció és d'emmagatzemar energia, aquests lípids comprenen el gruix del greix emmagatzemat en els teixits animals. L'hidròlisi dels enllaços èster dels triacilglicerols i l'alliberament de glicerol i d'àcids grassos del teixit adipós són els passos inicials en la metabolització del greix.[36]
Les subclasses addicionals de glicerolípids estan representades pels glucosilglicerols, els quals estan caracteritzats per la presència d'un o més monosacàrids units al glicerol via l'enllaç glucosídic. Exemples d'estructures d'aquesta categoria són els digalactosildiacilglicerols que es troben en les membranes de les plantes[12] i els seminolípids dels espermatozoides dels mamífers.[37]


Triacilglicerols
[modifica]

Els triacilglicerols estan constituïts per la unió mitjançant una reacció d'esterificació d'una molècula de glicerina (propanotriol) amb tres àcids grassos.
Amb una reacció d'hidròlisi podem alliberar els àcids grassos i la glicerina per tal de transformar-los en energia en forma d'ATP. Per aquest motiu, els triacilglicerols constitueixen la reserva energètica dels organismes vius.
Constitueixen la principal reserva energètica dels organismes animals (greixos) i vegetals (olis). L'excés de lípids és emmagatzemat en grans dipòsits en els animals en teixits adiposos. Són bons aïllants tèrmics que s'emmagatzemen en els teixits adiposos subcutanis dels animals de climes freds com per exemple, la balena, l'os polar, etc. Són productors de calor metabòlica, durant la seva degradació. Un gram de greix produeix 9,4 kcal. En les reaccions metabòliques d'oxidació (les proteïnes i glúcids produeixen 4,1 kcal). Donen protecció mecànica, com els constituents dels teixits adiposos que estan situats a la planta del peu, palmell de la mà i envoltant el ronyó (embolcallant-lo i evitant el seu despreniment).
Cèrids
[modifica]

Les ceres es formen per la unió d'una àcid gras de cadena llarga amb un alcohol, també de cadena llarga, mitjançant una reacció de saponificació, és a dir, són èsters d'àcids grassos. Els alcohols més habituals són el cetílic, C16H33OH; el carnaubílic, C24H49OH i el miricílic, C30H61OH. Són importants la cera carnauba obtinguda d'una espècie de palma; la cera de les abelles; la lanolina, que és la cera de la llana; l'espermaceti, que es troba al cap dels catxalots, etc.
La principal característica d'aquestes substàncies és la seva impermeabilitat, per la qual cosa els organismes vius l'empren per a impermeabilitzar els seus teixits de revestiment exterior o bé per a netejar-los. Una altra característica és que tenen un punt de fusió alt, sempre per sobre dels 60 °C. S'empren per a vernissos i cremes per a les sabates, per a fabricar candeles i espelmes, i en productes farmacèutics.
Fosfolípids
[modifica]

Els fosfolípids són cadascun dels lípids de les cèl·lules que per hidròlisi poden lliurar àcid fosfòric, H3PO4. Ocorren, quasi exclusivament, en les membranes cel·lulars i, en petites quantitats, en els greixos de dipòsit. El teixit nerviós n'és especialment ric.
Químicament són dièsters de l'àcid fosfòric, el qual esterifica, d'una banda, un dels alcohols (colina, etanolamina, serina, inositol, etc.), i de l'altra, la glicerina o bé l'esfingosina. Un grup molt gran i molt variat de fosfolípids és constituït pels fosfoglicèrids o glicerilfosfàtids, que són els fosfolípids que contenen glicerina. Les esfingomielines són fosfolípids que contenen esfingosina i que, per tant, formen part dels esfingolípids.[33]
Icosanoides
[modifica]
Icosanoide és el nom general que se li dona a un grup de molècules de constitució lipídica obtingudes de l'oxigenació dels àcids grassos essencials de 20 carbonis tipus omega-3 i omega-6. Compleixen àmplies funcions com mediadors per al sistema nerviós central, la inflamació i la resposta immune tant de vertebrats com invertebrats.
Tots els icosanoides són molècules de 20 àtoms de carboni i estan agrupats en prostaglandines, tromboxans, leucotriens, i certs hidroxiàcids precursors dels leucotriens. Constitueixen les molècules involucrades en les xarxes de comunicació cel·lular més complexes de l'organisme animal, incloent l'home.
Els icosanoides són àcids carboxílics, fet del qual deriven les seves propietats metabòliques.[38] Els derivats de l'omega-6 (ω-6), en general, tenen peculiaritats antiinflamatòries, no tant així els derivats de l'omega-3 (ω-3). Dels icosanoides, les prostaglandines i els tromboxans inclouen anells saturats i heterocíclics, mentre que els leucotriens i els hidroxiàcids són lineals (obertes de principi a fi de la seva cadena). Tots ells deriven de l'àcid araquidònic (AA), i són àcids grassos essencials. A més de l'àcid araquidònic, altres àcids grassos essencials també poden servir com precursors en la formació d'aquestes molècules, per exemple, l'àcid linoleic i l'àcid linolènic.
 |  |  | |
| Prostaglandina E1. L'anell de cinc costats és característic de la seva classe. | Tromboxà A2. Els oxígens s'han endinsat en l'anell. | Leucotriè B4. Presenta tres dobles enllaços conjugats. | |
 |  | ||
| Prostaciclina I2. El segon anell el distingeix de les prostaglandines. | Leucotriè E4, un exemple d'un leucotriè "cisteinil". | ||
- Les prostaglandines tenen 20 àtoms de carboni, un grup d'àcid carboxílic i un anell de cinc carbonis com a part de la seva estructura. Totes les prostaglandines tenen un ciclopentà (un anell de cinc (penta) carbonis), excepte la prostaglandina I2, que té un anell addicional.
- Els tromboxans són molècules cícliques (heterociclícliques) bé sigui de 6 carbonis o de 5 carbonis amb 1 oxigen, formant aquest últim un petit anell d'oxà —un anell que conté 5 àtoms de carboni i un àtom d'oxigen. Tenen estructures semblants a les prostaglandines i segueixen la mateixa nomenclatura. Consten d'un anell i dues cues. Es van trobar primerament en els trombòcits (plaquetes), d'allí el seu nom tromboxà.
- Els leucotriens són molècules lineals. Es van identificar en leucòcits i per això se'ls coneix com a leucotriens. Encara que tenen quatre enllaços dobles, inicialment es pensava que tenien 3 dobles enllaços conjugats (d'allí trieno). La seva producció en el cos forma part d'una complexa seqüència metabòlica que inclou la producció d'histamina.
Lípids insaponificables
[modifica]Els lípids insaponificables són aquells que no contenen àcids grassos. En conseqüència, no poden formar la reacció de saponificació. Dintre dels lípids insaponificables podem trobar els esteroides i els terpens.
Esteroides
[modifica]

Dit de cadascun dels components del grup de substàncies naturals que contenen l'esquelet fonamental del ciclopentanperhidrofenantrè, en el qual hi ha fixades una cadena lateral o unes quantes. Molts composts esteroides són deguts a l'existència de formes estereoisòmeres, a la presència eventual de dobles enllaços insaturats i a la introducció de distints substituents en els grups R1, R2, R3 als carbonis 10, 12 i 17 respectivament de la figura adjunta; així, en el cas dels esteroides naturals, els substituents en R1 i R2 són sempre grups metílics, i en el cas de l'estrofantidina i l'aldosterona, R1 i R2 són grups aldehídics. El nucli esteroide és format pels tres cicles del ciclohexà i un cicle de ciclopentà. Els sistemes del ciclohexà adopten la forma més estable de cadira. En fisiologia humana i medicina els esteroides més importants són el colesterol, les hormones esteroides, els seus precursors i els seus metabòlits.





Terpenoides
[modifica]


Els terpens o isoprenoides són polimeritzacions de l'isoprè. Hom els sol classificar, anàlogament als terpens, en hemiterpenoides, monoterpenoides, sesquiterpenoides, etc., d'acord amb el nombre d'àtoms de carboni de llurs estructures.
La distribució natural dels diferents tipus de terpenoides és irregular.
- Els monoterpenoides i sesquiterpenoides ocorren com a constituents principals dels olis essencials de nombroses plantes i són molt àmpliament difosos en la natura. També moltes espècies d'insectes secreten terpenoides d'aquests tipus com a hormones i feromones. Entre aquests hom pot esmentar:
- Els diterpenoides, tot i ésser menys abundants, es troben també molt difosos. Així, el fitol ocorre en les plantes verdes com a constituent de la clorofil·la, mentre que l'àcid abiètic és el component més important de les oleoresines exsudades per les coníferes.
- Els triterpenoides ocorren principalment en plantes, tant com a constituents com a la saba, en forma lliure o com a èsters i glicòsids. No obstant això, alguns triterpenoides, com l'esqualè i el lanosterol, ocorren en les espècies animals com a precursors biosintètics dels esteroides.
- Pel que fa als tetraterpenoides, els de més importància són els carotenoides. El carotè està format per la polimerització de vuit isoprens. Proporciona una coloració de color vermell. Present al tomàquet i la pastanaga, a partir d'ella el fetge és capaç de sintetitzar dues molècules de vitamina A (polimerització de quatre isoprens), que prevé la ceguesa crepuscular. La xantofil·la també està constituïda per la polimerització de vuit isoprens, amb la peculialitat que té un grup alcohol a cada extrem. Això fa que proporcioni una coloració de color groc. Està present en algunes flors, com la margarida.
- Finalment, els terpenoides de natura polimèrica (politerpenoides), com el cautxú i la gutaperxa, existeixen en un nombre reduït d'espècies vegetals.
Els terpenoides tenen aplicació, tant en forma pura com en barreges, en farmàcia, alimentació i perfumeria, com a dissolvents, penetrants, agents mullants i conservants, i tot el grup és d'una gran importància econòmica. D'altra banda, l'estudi dels terpenoides pel que fa a llur separació, elucidació estructural, síntesi i reactivitat, ha estat un dels motors del desenvolupament de la química orgànica moderna.[33]
-

-

-

-

-
-
 β-carotè
β-carotè




Catabolisme
[modifica]
El catabolisme és el conjunt de processos metabòlics que alliberen energia i inclouen degradació i oxidació de les molècules d'aliment. El propòsit d'aquestes reaccions és aconseguir energia i components necessaris per a les reaccions anabòliques, de biosíntesi de noves molècules.
Els greixos són catalitzats per uns enzims anomenats lipases, que realitzen la hidròlisi a àcids grassos i glicerol. El glicerol entra en la glucòlisi i els àcids grassos són degradats per beta oxidació per a alliberar acetil-CoA, que després passa al cicle de Krebs. Degut a les elevades proporcions del grup metilè, els àcids grassos alliberan més energia en la seva oxidació que els glúcids, ja que els glúcids com la glucosa tenen més oxigen en les seves estructures.
Anabolisme
[modifica]
L'anabolisme és el conjunt de processos metabòlics constructius on l'energia alliberada pel catabolisme és utilitzada per a sintetitzar molècules complexes. En general, les molècules complexes que donen lloc a estructures cel·lulars són construïdes a partir de precursors simples.
Els àcids grassos se sintentizen al polimeritzar i reduir unitats d'acetil-CoA. Les cadenes en els àcids grassos són esteses per un cicle de reaccions que agreguen el grup acetil, ho reduïxen a alcohol, deshidraten a un grup alquè i després ho reduïxen novament a un grup alcà. Els enzims de la síntesi d'àcids grassos es dividixen en dos grups: en els animals i fongs, les reaccions de la síntesi són dutes a terme per una sola proteïna multifuncional tipus I, mentre que en plantes i bacteris són els enzims tipus II per separat les que duen a terme cada etapa en la ruta.
Els terpens i isoprenoides són sintentizats per la unió i modificació d'unitats d'isoprè donades pels precursors reactius pirofosfosfat d'isopentenil i pirofosfat de dimetilalil. Aquests precursors poden sintentizar-se de diversos modes. En animals se sintentitzen a partir d'acetil-CoA, mentre que en plantes i bacteris es fa a partir de piruvat i gliceraldehíd 3-fosfat com a substrats. Una reacció que usa aquests donadors isoprènics activats és la biosíntesi d'esteroides. En aquest cas, les unitats d'isoprenoides són unides covalentemente per a formar escualè, que es plega formant una sèrie d'anells donant lloc a una molècula denominada lanosterol. El lanosterol pot després ser transformat en esteroides com el colesterol.
Referències
[modifica]- ↑ Berg, J.M.; i col.. Bioquímica. Reverté, 2008. ISBN 8429176004.
- ↑ Fahy E, Subramaniam S., Brown H., Glass C., Merrill A., Murphy R., Raetz C., Russell D., Seyama Y., Shaw W., Shimizu T., Spener F., van Meer G., VanNieuwenhze M., White S., Witztum J., Dennis E. «A comprehensive classification system for lipids». J Lipid Res, 46, 5, 2005, pàg. 839–61. Arxivat de l'original el 2010-08-24. DOI: 10.1194/jlr.E400004-JLR200. PMID: 15722563 [Consulta: 12 juny 2009]. Arxivat 2010-08-24 a Wayback Machine.
- ↑ Enciclopèdia Catalana. Gran Diccionari de la llengua catalana, 1999.[Enllaç no actiu]
- ↑ 4,0 4,1 Domènech, X.; i col.. Bioelements i Biomolècules. Edicions Universitat Barcelona, 1992. ISBN 8475338968.
- ↑ 5,0 5,1 5,2 Fahy E, Subramaniam S, Murphy R, Nishijima M, Raetz C, Shimizu T, Spener F, Van Meer G, Wakelam M and Dennis E.A «Update of the LIPID MAPS comprehensive classification system for lipids». Journal of Lipid Research, 50, 2009, pàg. S9–S14. DOI: 10.1194/jlr.R800095-JLR200. PMID: 19098281.
- ↑ Subramaniam S, Fahy E, Gupta S, Sud M, Byrnes R.W, Cotter D, Dinasarapu A.R and Maurya M.R «Bioinformatics and Systems Biology of the Lipidome». Chemical Reviews, 111, 10, 2011, pàg. 6452–6490. DOI: 10.1021/cr200295k. PMID: 21939287.
- ↑ Michelle A, Hopkins J, McLaughlin CW, Johnson S, Warner MQ, LaHart D, Wright JD. Human Biology and Health. Englewood Cliffs, New Jersey, USA: Prentice Hall, 1993. ISBN 0-13-981176-1. OCLC 32308337.
- ↑ Brasaemle DL «Thematic review series: adipocyte biology. The perilipin family of structural lipid droplet proteins: stabilization of lipid droplets and control of lipolysis». J. Lipid Res, 48, 12, December 2007, pàg. 2547–59. Arxivat de l'original el 2010-07-06. DOI: 10.1194/jlr.R700014-JLR200. PMID: 17878492 [Consulta: 13 febrer 2012]. Arxivat 2010-07-06 a Wayback Machine.
- ↑ Stryer et al., p. 619.
- ↑ Stryer et al., pp. 329–331
- ↑ Heinz E.(1996). Plant glycolipids: structure, isolation and analysis. in Advances in Lipid Methodology - 3, pp. 211–332 (ed. W.W. Christie, Oily Press, Dundee)
- ↑ 12,0 12,1 Hölzl G, Dörmann P «Structure and function of glycoglycerolipids in plants and bacteria». Progress in Lipid Research, 46, 5, 2007, pàg. 225–43. DOI: 10.1016/j.plipres.2007.05.001. PMID: 17599463.
- ↑ Stryer et al., pp. 333–34.
- ↑ van Meer G, Voelker DR, Feigenson GW «Membrane lipids: where they are and how they behave». Nature Reviews Molecular Cell Biology, 9, 2, 2008, pàg. 112–24. DOI: 10.1038/nrm2330. PMC: 2642958. PMID: 18216768.
- ↑ Feigenson GW «Phase behavior of lipid mixtures». Nature Chemical Biology, 2, 11, 2006, pàg. 560–63. DOI: 10.1038/nchembio1106-560. PMC: 2685072. PMID: 17051225.
- ↑ Wiggins PM «Role of water in some biological processes». Microbiological Reviews, 54, 4, 1990, pàg. 432–49. PMC: 372788. PMID: 2087221.
- ↑ Raschke TM, Levitt M «Nonpolar solutes enhance water structure within hydration shells while reducing interactions between them». Proceedings of the National Academy of Sciences U.S.A, 102, 19, 2005, pàg. 6777–82. DOI: 10.1073/pnas.0500225102. PMC: 1100774. PMID: 15867152.
- ↑ Wang X «Lipid signaling». Current Opinions in Plant Biology, 7, 3, 2004, pàg. 329–36. DOI: 10.1016/j.pbi.2004.03.012. PMID: 15134755.
- ↑ Eyster KM «The membrane and lipids as integral participants in signal transduction». Advances in Physiology Education, 31, 1, 2007, pàg. 5–16. DOI: 10.1152/advan.00088.2006. PMID: 17327576.
- ↑ Hinkovska-Galcheva V, VanWay SM, Shanley TP, Kunkel RG «The role of sphingosine-1-phosphate and ceramide-1-phosphate in calcium homeostasis». Current Opinion in Investigational Drugs, 9, 11, 2008, pàg. 1192–205. PMID: 18951299.
- ↑ Saddoughi SA, Song P, Ogretmen B «Roles of bioactive sphingolipids in cancer biology and therapeutics». Subcellular Biochemistry, 49, 2008, pàg. 413–40. DOI: 10.1007/978-1-4020-8831-5_16. PMC: 2636716. PMID: 18751921.
- ↑ Klein C, Malviya AN «Mechanism of nuclear calcium signaling by inositol 1,4,5-trisphosphate produced in the nucleus, nuclear located protein kinase C and cyclic AMP-dependent protein kinase». Frontiers in Bioscience, 13, 2008, pàg. 1206–26. DOI: 10.2741/2756. PMID: 17981624.
- ↑ Boyce JA «Eicosanoids in asthma, allergic inflammation, and host defense». Current Molecular Medicine, 8, 5, 2008, pàg. 335–49. DOI: 10.2174/156652408785160989. PMID: 18691060.[Enllaç no actiu]
- ↑ Bełtowski J «Liver X receptors (LXR) as therapeutic targets in dyslipidemia». Cardiovascular Therapy, 26, 4, 2008, pàg. 297–316. DOI: 10.1111/j.1755-5922.2008.00062.x. PMID: 19035881.
- ↑ Indiveri C, Tonazzi A, Palmieri F «Characterization of the unidirectional transport of carnitine catalyzed by the reconstituted carnitine carrier from rat liver mitochondria». Biochim. Biophys. Acta, 1069, 1, October 1991, pàg. 110–6. DOI: 10.1016/0005-2736(91)90110-T. PMID: 1932043.
- ↑ Parodi AJ, Leloir LF «The role of lipid intermediates in the glycosylation of proteins in the eucaryotic cell». Biochim. Biophys. Acta, 559, 1, April 1979, pàg. 1–37. DOI: 10.1016/0304-4157(79)90006-6. PMID: 375981.
- ↑ Helenius A, Aebi M «Intracellular functions of N-linked glycans». Science, 291, 5512, 2001, pàg. 2364–69. DOI: 10.1126/science.291.5512.2364. PMID: 11269317.
- ↑ M. Nowicki and M. Frentzen «Cardiolipin synthase of Arabidopsis thaliana». FEBS Letters, 579, 10, 2005, pàg. 2161–2165. DOI: 10.1016/j.febslet.2005.03.007. PMID: 15811335.
- ↑ M. Nowicki «Characterization of the Cardiolipin Synthase from Arabidopsis thaliana». Ph.D. thesis, RWTH-Aachen University, 2006. Arxivat de l'original el 2011-10-05 [Consulta: 13 febrer 2012].
- ↑ Gohil VM, Greenberg ML «Mitochondrial membrane biogenesis: phospholipids and proteins go hand in hand». Journal of Cell Biology, 184, 4, 2009, pàg. 469–72. DOI: 10.1083/jcb.200901127. PMC: 2654137. PMID: 19237595.
- ↑ Hoch FL «Cardiolipins and biomembrane function». Biochimica et Biophysica Acta, 1113, 1, 1992, pàg. 71–133. PMID: 10206472.
- ↑ «Virtual ChemBook» (en anglès). Arxivat de l'original el 2015-03-15. [Consulta: 27 desembre 2020].
- ↑ 33,0 33,1 33,2 «Lípid». Gran Enciclopèdia Catalana. Barcelona: Grup Enciclopèdia Catalana.
- ↑ Marcel Mayol. Fonaments i Tècniques d'Anàlisi Bioquímica.
- ↑ Coleman RA, Lee DP «Enzymes of triglyceride synthesis and their regulation». Progress in Lipid Research, 43, 2, 2004, pàg. 134–76. DOI: 10.1016/S0163-7827(03)00051-1. PMID: 14654091.
- ↑ van Holde and Mathews, p. 630–31.
- ↑ Honke K, Zhang Y, Cheng X, Kotani N, Taniguchi N «Biological roles of sulfoglycolipids and pathophysiology of their deficiency». Glycoconjugates Journal, 21, 1–2, 2004, pàg. 59–62. DOI: 10.1023/B:GLYC.0000043749.06556.3d. PMID: 15467400.
- ↑ Consejo General de Colegios Oficiales de Farmacéuticos
 PDF
PDF
Bibliografia
[modifica]- Akoh, C.C.; Min, D.B. Food lipids: chemistry, nutrition, and biotechnology (en anglès). 2a ed.. CRC Press, 2002. ISBN 0824707494.
- Betteridge, J. Lipids: current perspectives (en anglès). Taylor & Francis, 1996. ISBN 1853172316.
- Donatelle, R. J. Health, The Basics. 6th ed. San Francisco: Pearson Education, Inc. 2005.
- Mozaffarian D, Katan MB, Ascherio A, Stampfer MJ, Willett WC (April 2006). "Trans Fatty Acids and Cardiovascular Disease". New England Journal of Medicine 354 (15):1601-1613. PMID: 16611951]
- «Nomenclature of Lipids». IUPAC-IUB Commission on Biochemical Nomenclature (CBN). [Consulta: 12 juny 2009].
- Spiller, G.A. Handbook of lipids in human nutrition (en anglès). CRC Press, 1996. ISBN 0849342481.
- Tyman, J.H.P. Lipids in health and nutrition (en anglès). Woodhead Publishing, 1999. ISBN 0854047980.
- Vance, D.E.; Vance, J.E. Biochemistry of Lipids, Lipoproteins and Membranes (en anglès). 5a edició. Elsevier, 2008. ISBN 0444532196.
| Principals famíles bioquímiques | ||
| Àcids nucleics | Alcaloides | Aminoàcids | Carbohidrats | Carotenoides | Cofactors enzimàtics | Esteroides | Flavonoides | Glicòsids | Lípids | Pèptids | Policètids | Tetrapirrols | Terpens | ||
| Anàlegs d'àcids nucleics: | Anàlegs d'àcids nucleics : | |
| Conceptes generals | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Rutes específiques |
| ||||||||||||||||||