Smjena generacija


Smjena generacija – poznata i kao smjena faza i metageneza – je tip životnog ciklusa koji se javlja kod nekih biljaka i algi u grupama Archaeplastida i Heterokontophyta koje imaju izražene haploidne spolne i diploidne bespolne faze. U tim skupinama, višećelijski gametofit, koji ima haploidan (n) broj hromosoma, smjenjuje se s višećelijskim sporofitom, koji je diploidnim 2n brojem hromosoma, sačinjenog od "n"parova. Zreli sporofit stvara spore, putem mejoze, u procesu kojim se broj hromosoma smanjuje na pola, iz 2n na n.[1][2]
Haploidne spore klijaju i izrastu u haploidni gametofit. U zrelosti, gametofit stvara gamete putem mitoze, koja ne menja broj hromosoma. Dva gameta (koji potiču iz različitih organizama iste vrste ili iz istog organizma) oplodnjom stvaraju zigot, koji se razvija u diploidni sporofit. Ovaj ciklus, od gametofita do gametofita (ili podjednako od sporofita do sporofita), način je na koji se sve kopnene biljke i mnoge alge podvrgavaju spolnoj reprodukciji.
Odnos sporofita i gametofita varira između različitih grupa biljaka. U onim algama koje imaju naizmjenično generacije, sporofiti i gametofiti su zasebni – neovisni organizmi koji mogu ali ne moraju imati sličan izgled. U mahovina jetrenjača i Anthocerotophyta, sporofit je manje razvijen od gametofita i u velikoj mjeri ovisi o njemu. Iako sporofiti mahovine mogu fotosintetizirati, potrebna je dodatna fotosinteza iz gametofita kako bi se održao rast i razvoj spora i ovisili o njemu za opskrbu vodom, mineralnim hranjivim tvarima i dušikom. Nasuprot tome, u svim modernim vaskularnim biljkama gametofit je manje razvijen od sporofita, iako su njihovi devonski preci imali gametofite i sporofite približno jednake složenosti. U paprati, gametofit je mali i spljošten autotrofni protalus na kojem je mladi sporofit kratkotrajno ovisan u svojoj prehrani. U cvjetnica je smanjenje gametofita mnogo ekstremnije; sastoji se od samo nekoliko ćelija koje u potpunosti rastu unutar sporofita.
Životinje se drugačije razvijaju. Oni direktno proizvode haploidne gamete. Ne nastaju haploidne spore koje bi se mogle podijeliti, tako da nemaju haploidni gametofit koji se izmjenjuje s diploidnim sporofitom. (Neki insekti imaju poseban sistem određivanja spola, pri čemu se haploidni mužjaci proizvode od neoplođenih jajašaca; međutim, ženke su diploidne), Životni ciklusi biljaka i algi s naizmjeničnim haploidnim i diploidnim višećelijskim stadijima nazivaju se "'diplohaplontski'" (ekvivalentni pojmovi haplodiplontski, diplobiontski ili dibiontski također su u upotrebi). Životni ciklusi, kao što su kod životinja u kojima postoji samo diploidna višećelijska faza, nazivaju se "'diplontski'" (diplont). Životni ciklusi u kojima postoji samo haploidna višećelijska faza nazivaju se "'haplontski'" (haplont)
Definicija
[uredi | uredi izvor]Smjena generacija definira se kao smjena višećelijskih diploidnih i haploidnih oblika u životnom ciklusu organizma, bez obzira jesu li ti oblici slobodni i ne.[3] Kod nekih vrsta, kao što je alga Ulva lactuca, diploidni i haploidni oblici doista su neovisni organizmi koji žive slobodno, uglavnom jednaki po izgledu i stoga se kaže da su izomorfni. Slobodno plivajući haploidni gameti tvore diploidni zigot koji klija u višećelijski diploidni sporofit. Sporofit stvara slobodno plivajuće haploidne spore koje klijaju u haploidne gametofite.[4] Međutim, u nekim drugim skupinama, ili sporofit ili gametofit je vrlo smanjen i nije sposoban za slobodno življenje. Naprimjer, u svim Bryophyta generacija gametofita je dominantna, a sporofit ovisi o njemu. Suprotno tome, kod svih modernih vaskularnih kopnenih biljaka gametofiti su jako reducirani, iako fosilni dokazi upućuju na to da su izvedeni od izomorfnih predaka. U biljka sjemenjača , ženski gametofit se potpuno razvija unutar sporofita koji ga štiti i njeguje, te embriona sporofita koji proizvodi. Zrno polena, koje je muški gametofiti, svodi se na samo nekoliko ćelija (po samo tri u mnogim slučajevima). Ovdje je pojam dvije generacije manje očit; kao što kažu Bateman & Dimichele: "(s)porofiti i gametofiti učinkovito funkcioniraju kao jedinstveni organizam".[5] The alternative term 'alternation of phases' may then be more appropriate.[6]
Historija
[uredi | uredi izvor]Rasprave o smjeni generacija u ranom dvadesetom stoljeću mogu biti zbunjujuće jer postoje različiti načini klasificiranja „generacija“ (seksualni nasuprot aseksualnom, gametofiti nasuprot sporofita, haploidni – diploidni itd.).[7] U početku su Chamisso i Steenstrup opisali sukcesiju različito organizovanih generacija (seksualnih i aseksualnih) kod životinja kao "naizmjeničenje generacija", proučavajući razvoj životinja Tunicata , Cnidaria i Trematoda životinje. Ovaj fenomen je takođe poznat kao heterogamija. Sada se pojam "smjena generacija" gotovo isključivo povezuje sa životnim ciklusima biljaka, tačnije s izmjenom haploidnih gametofita i diploidnih sporofita.[7]
Wilhelm Hofmeister pokazao je morfološku smjenu generacija kod biljska,[2] između generacije koja nosi spore (sporofit) i generacije koja podnosi gamete (gametofit).[8][9][10] U to se vrijeme otvorila rasprava koja se usredotočila na porijeklo aseksualne generacije kopnenih biljaka (tj. Sporofita) i konvencijski je okarakterizirana kao sukob između teorija: antiteza (Čelakovský, 1874.) i homologije (Pringsheim, 1876.) smjena generacija.[7] Prema nekima, Čelakovský je skovao riječi sporofit igametofit.
Eduard Strasburger (1874) otkrio je smjenu generacija između diploidne i haploidne faze jedra,[7] zvanu citološka smjena jedrovih faza.[11] Iako se najčešće podudaraju morfološka smjena i smjena jedarnih faza ponekad su neovisne jedna o drugoj, npr. kod mnogih crvenih algi, ista nukleusna faza može odgovarati dvjema različitim morfološkim generacijama. Kod nekih paprati koje su izgubile seksualnu reprodukciju, u nukleusnoj fazi nema promjene, ali smjena generacija se održava.[12]
Smjena generacija kod biljaka
[uredi | uredi izvor]Osnovni elementi
[uredi | uredi izvor]Gornji dijagram prikazuje osnovne elemente smjene generacijakod biljaka. Mnoge varijacije koje su pronađene u različitim skupinama biljaka opisane su korištenjem ovih koncepata kasnije u članku. Polazeći s desne strane dijagrama, uključeni su sljedeći procesi:
- Dva jednoćelijska haploidna gameta, od kojih svaki sadrži neuparene hromosome n, spajaju se tako da formiraju jednoćelijski diploidni zigot, koja sada sadrži n parova hromosoma, odnosno ukupno 2n.
- Jednoćelijski diploidni zigot klija, dijelivši se normalnim procesom (mitoze), koji održava broj hromosoma na 2n. Rezultat je multićelijski diploidni organizam, nazvan sporofit (jer u zrelosti proizvodi spore).
- Kada dostigne zrelost, sporofit stvara jednu ili više sporangija, koje su organi koji proizvode diploidne matične ćelije spore (sporociti). Oni se dijele posebnim postupkom (mejoza) koji smanjuje broj hromosoma za polovinu. To u početku rezultira u četiri jednoćelijske haploidne spore, a svaka sadrži n neuparenih hromosoma.
- Jednoćelijska haploidna spora klija, dijelivši se normalnim procesom (mitozom), koji održava broj hromosoma na n. Rezultat je multićelijski haploidni organizam, nazvan gametofit (jer u zrelosti stvara gamete).
- Kada dostigne zrelost, gametofit stvara jednu ili više gametangija, koje su organi za proizvodnju haploidnih gameta. Barem jedna vrsta gameta posjeduje neki mehanizam za dostizanje drugog gameta da bi se s njim spojila.
„Smjena generacija“ u životnom ciklusu je tako između diploidne (2n) generacije sporofita i haploidne (n) generacije gametofita.

Situacija je prilično različita od one kod životinja, gdje je temeljni proces da diploidna (2n) jedinka direktno stvara haploidne (n) gamete mejozom. Spore (tj. haploidne ćelije koje su u stanju da se podvrgnu mitozi) ne nastaju, tako da nema ni aseksualne multićelijske generacije koja se smjenjuje sa seksualnom višećelijskom generacijom. (Neki insekti – kao pčele, naprimjer, imaju haploidne mužjake koji se razvijaju iz neplođenih jajašaca, ali sve su ženke diploidne.)
Varijacije
[uredi | uredi izvor]Dijagram koji je gore prikazan dobar je prikaz životnog ciklusa nekih višećelijskih algi (npr. rod Cladophora), koji imaju sporofite i gametofite gotovo identičnog izgleda i koji nemaju različite vrste spora ili gameta.[13] Međutim, postoji mnogo mogućih varijacija temeljnih elemenata životnog ciklusa koji imaju smjenu generacija. Svaka varijacija može se pojaviti odvojeno ili u kombinaciji, što rezultira čudesnom raznolikošću životnih ciklusa. Izrazi koje koriste botaničari u opisivanju ovih životnih ciklusa mogu biti podjednako zbunjujući. Kao što kažu Bateman i Dimichele "[...] smjena generacija je postala terminološka mora; često jedan pojam predstavlja nekoliko pojmova ili je jedan pojam predstavljen s više pojmova“.[14] Moguće varijacije su:
- Relativni značaj sporofita i gametofita
- Jednake (homomorfija ili izomorfija).
Končaste alge iz roda' 'Cladophora' , koje se uglavnom nalaze u svježem vodama, imaju diploidne sporofite i haploidne gametofite koji su spolja nerazlučivi. Evolucija smjene generacija sugerira da su to činili njihovi preci.[15] Nijedna današnja kopnena biljka nema podjednako dominantne sporofite i gametofite, mada neke teorije evolucije smjene generacija sugeriraju da su to činile i kopnene predačke biljke. - Nejednako (heteromorfija ili anizomorfija)

Gametofit mahovine Mnium hornum - Dominantni gametofit (gametofitne).
Kod jetrenjača, mahovina i paprati, dominantni oblik je haploidni gametofit. Diploidni sporofit nije sposoban za samostalan opstanak, rastući pretežnood roditeljskog gametofita, a hlorofil dobija tek kada sazrije.[16]
Sporofit paprati Blechnum discolor - Dominantni sporofit (sporofitne).
U paprati su sporofit i gametofit sposobni za samostalan život, ali dominantan oblik je diploidni sporofit. Haploidni gametofit je mnogo manji i jednostavnije građe. U biljkama sjemenjačama, gametofit se još više smanjuje (najmanji, na samo tri ćelije), svu svoju hranu dobija od sporofita. Ekstremno smanjenje veličine gametofita i njegovo zadržavanje unutar sporofita znači da se, kada se primjenjuje na sjemenskim biljkama, izraz „smjena generacija“ pomalo dovodi u zabludu: "[s]porofit i gametofit efikasno funkcioniraju kao jedinstven organizam".[5] Neki autori preferirali su termin "smjena faza".[6]
- Dominantni gametofit (gametofitne).
- Jednake (homomorfija ili izomorfija).
- Diferencijacija gameta
- Oba gameta su jednaka (izogamija).
Kao i druge vrste roda Cladophora, C. callicoma ima bičaste gamete koji su identičnog izgleda i sposobnosti kretanja.[15] - Gameti različite veličine (anizogamija).
- Oba gameta slične pokretljivosti.
Vrste morske salate Ulva imaju gamete sa po dva biča za kretanje. Oni su, međutim, različite veličine: veći su ženski, a manji muški.[17] - Jedan veliki i sesilan, drugi malen i pokretan (oogamija). Veći sjedeći megagameti su jajašca (jajne ćelije), a manji pokretni mikrogameti su spermatozoidi. Stepen pokretljivosti spermija može biti vrlo ograničen (kao u slučaju cvjetnica), ali svi se mogu kretati prema sjedećim jajima. Kada se (kao što je gotovo uvijek slučaj) stvaraju spermatozoidi i jajašca u različitim vrstama gametangia, one koje stvaraju spermu nazivaju se antheridija (anteridij), a one koje stvaraju jajašca arhegonija (arhegonij).

Gametofit Pellia epiphylla sa sporofitima koji rastu iz ostataka arhegonije. - Anteridij i arhegonij javljaju se na istom gametofitu, zvanom monoični. (Mnogi izvori, uključujući one koji se bave mahovinama, koriste izraz "monoecijski" za ovu situaciju, a "diecijski" za suprotno.[18][19] Ovde se „monoecijski“ i „diecijski“ upotrebljavaju samo za sporofite.)
Jetrenjača Pellia epiphylla ima gametofit kao dominantnu generaciju. Ona je monoecijska: mala anteridija koja proizvodi crvenkastu spermu raspršena je po sredini, dok arhegonija (koja proizvodi jaja) raste bliže vrhovima podjela biljke.[20] - Anteridij i arhegonij pojavljuju se na različitim gametofitima, koji su diecijski.
Mahovina Mnium hornum ima gametofit kao dominantnu generaciju. Ona je diecijska: muške biljke stvaraju samo anteridije u vršnim rozetama, a ženske – samo arhegonije u obliku stabljikastih kapsula.[21] Sjeme biljnih gametofita je također dvoslojno. Međutim, roditeljski sporofit može biti jednoličan, proizvodeći muške i ženske gametofite ili diecijski, proizvodeći gametofite samo jednog spola. Biljke sjemenjače imaju gametofit izuzetno smanjene veličine; arhegonij se sastoji od samo malog broja ćelija, a cio muški gametofit može biti predstavljen na samo dve ćelije.
- Anteridij i arhegonij javljaju se na istom gametofitu, zvanom monoični. (Mnogi izvori, uključujući one koji se bave mahovinama, koriste izraz "monoecijski" za ovu situaciju, a "diecijski" za suprotno.[18][19] Ovde se „monoecijski“ i „diecijski“ upotrebljavaju samo za sporofite.)
- Oba gameta slične pokretljivosti.
- Oba gameta su jednaka (izogamija).
- Diferencijacija spora
- Sve spore iste veličine (homosporija ili izosporija). ).
konjski repovi (vrste roda Equisetum) imaju spore koje su sve iste veličine. - Dvije spore različite veličine (heterosporija ili anizosporija): veće megaspore i manje mikrospore. Kad se dvije vrste spora proizvode u različitim vrstama sporangija, one se nazivaju megasporangije i mikrosporangije. Megaspora se često (ali ne uvijek) razvija na štetu ostalih triju ćelija proisteklih iz mejoze, koje abortiraju.
- Sve spore iste veličine (homosporija ili izosporija). ).



Gametofiti sjemenjača također su diecijki. Međutim, roditeljski sporofit može biti jednoličan, proizvodeći muške i ženske gametofite ili dvodomi, proizvodeći gametofite samo jednog spola. Njihovi ametofiti su izuzetno smanjene veličine; arhegonij se sastoji od malog broja ćelija, a cio muški gametofit može biti predstavljen sa samo dvije ćelije.
- Megasporangija i mikrosporangija javljaju se na istom sporofitu, koji se tada zove monoecijski.
Većina cvjetnica spada u ovu kategoriju. Tako cvijet ljiljana sadrži šest stabljika (mikrosporangije) koje stvaraju mikrospore; one se razvijaju u polenovo zrno (mikrogametofit) i tri spojena karpela koji stvaraju integrirane megasporangije (ovule) od kojih svaki stvara megasporu. Ona se razvija u megasporangiju, da bi proizvela megagametofit. U drugim biljkama, kao što je lješnik, neki cvjetovi imaju samo stabljike, druge samo prašnike, ali ista biljka (tj. porofit) ima obje vrste cvijeta i tako je monoecijska.
Cvjetovi evropske božikovine, dvodome vrste: muška iznad, ženska dolje (lišće odrezano da bi se jasnije prikazali cvjetovi) - Megasporangija i mikrosporangija na različitim sporofitima koji su, dakle 'diecijski (dvodomi).
Pojedinačna stabla evropske boćikovine ( Ilex aquifolium ) proizvode ili 'muške' cvjetove koji imaju samo funkcionalne stabljike (mikrosporangije), proizvodeći mikrospore koje se razvijaju u polenovo zrno (mikrogametofiti) ili 'ženski' cvjetovi koje ima samo funkcionalnu karpelu koja proizvodi integrirane megasporangije (ovule); one sadrže megaspore koje se razvija u višećelijski megagametofit.
- Megasporangija i mikrosporangija javljaju se na istom sporofitu, koji se tada zove monoecijski.

Između tih varijacija postoje neke korelacije, ali one su upravo to, korelacije, a ne i apsolutni odnosi. Naprimjer, kod cvjetnica mikrospore na kraju proizvode mikrogamete (spermu), a megaspore –megagamete (jajašca). Međutim, kod paprati i njihovih srodnika postoje grupe sa nediferenciranim sporama, ali diferenciranim gametofitima. Naprimjer, paprat Ceratopteris talictrioidi ima spore samo jedne vrste, koje se neprekidno razlikuju po veličini. Manje spore imaju tendenciju klijanja u gametofite koji proizvode samo antheridij koji proizvodi spermu.
Kompleksni životni ciklus
[uredi | uredi izvor]
Dijagram prikazuje smjenu generacijakod vrste koja je heteromorfna, sporofitska, oogametska, diecijska, heterosporna i dvoprirodna. Primjer sjemenjače može biti stablašica vrba (većina vrsta roda Salix je dvospolna).[22] Polazeći od središta dijagrama, uključeni procesi su:
- Nepomično jaje, sadržano u arhegoniju, stapa se s pokretnom spermom, oslobođenom antheridija. Rezultirajući zigot je ili muški ili ženski.
- Muški zigot razvija se mitozom u mikrosporofit koji u zrelosti proizvodi jednu ili više mikrosporangija. Mikrospore se mejozom razvijaju u mikrosporangiju.
Kod vrbe (poput svih sjemenjača) zigot se najprije razvija u mikrosporofit embriona, unutar ovule (megasporangija zatvorena u jedan ili više zaštitnih slojeva tkiva poznatih kao integument). Sazrevanjem ove strukture postaje seme. Kasnije se sjeme rasipa, klija i izraste u zrelo drvo. „Muško“ stablo vrbe (mikrosporofit) proizvodi cvjetove sa samo pršnicima, od kojih su antere mikrosporangije. - Mikrospore klijaju proizvodeći mikrogametofite; u zrelosti nastaje jedna ili više antheridija. Spermatozoidi se razvijaju u antheridiji.
Kod vrbe, mikrospore se ne oslobađaju od antera (mikrosporangija), već se razvijaju u polenova zrna (mikrogametofiti) unutar nje. Rasijava se cjelokupno zrno polena (npr. insektima ili vjetrom) u ovule (megagametofit), gdje se stvara sperma koja se kreće niz polenovu cijev do jajne ćelije. - Ženski zigot razvija se putem mitoze u megasporofit, koji kad sazrije, proizvodi jednu ili više megasporangija. Megaspore se razvijaju unutar megasporangije; tipski, jedna od četiri spore proizvedene mejozom razvijaju se na račun preostale tri, koje nestaju.
'Ženske' vrbe (megasporofiti) proizvode cvjetove samo sa tučkovima (modificirano lišće koje nosi megasporangiju). - Megaspore klijaju stvarajući megagametofite; u zrelosti se proizvodi jedna ili više arhegonija. Jaja se razvijaju u arhegoniji.
Karpele vrbe proizvode ovule, megasporangije su zatvorene u integumente. Unutar svake ovule mitozom se razvija megaspora u megagametofit. Arhegonij se razvija unutar megagametofita i stvara jaje. Čitava „gametofitska generacija“ ostaje pod zaštitom sporofita, osim polenovih zrnaca (svedena u tri ćelije koje se nalaze u zidu mikrospore).
- Muški zigot razvija se mitozom u mikrosporofit koji u zrelosti proizvodi jednu ili više mikrosporangija. Mikrospore se mejozom razvijaju u mikrosporangiju.
Životni ciklusu po grupama biljaka
[uredi | uredi izvor]Termin "biljke" ovdje uključuje Archaeplastida, tj. Glaucophyta , crvene i zelene alge i kopnene biljke.
Smjena generacija se javlja u gotovo svim višećelijskim crvenim i zelenim algama, kod slatkovodnih formi (kao što je Cladophora) i morskih (kao što su u rodu Ulva ). U većini generacije su homomorfne (izomorfne) i slobodno žive. Neke vrste crvenih algi imaju složenu trofaznu smjenu generacija u kojoj postoji faza gametofita i dvije različite sporofitne faze. Za dodatne informacije pogledajte Crvene alge: Reprodukcija.
Sve kopnene biljke imaju heteromorfnu (anizomorfnu) smjenu generacija, u kojoj se sporofiti i gametofiti izrazito razlikuju. Kod svih mahovina, tj. Marchantiophyta i paprati, generacija gametofita se najočitije ispoljavaju. Kao ilustraciju razmotrite jednoličnu mahovinu. Antheridij i arhegonij razvijaju se na zreloj biljci (gametofit). U prisustvu vode, biflagelatni spermatozoidi iz antheridije doplivaju do arhegonije i dolazi do oplodnje, što dovodi do stvaranja diploidnog sporofita. Sporofiti rastu iz arhegonija. Njihovo tijelo sastoji se od duge stabljike na vrhu kapsule unutar koje ćelije koje proizvode spore prolaze kroz mejozu i formiraju haploidne spore. Većina mahovina spore raspršuje vjetrom, iako Splachnum sphaericum entomophilous, privlači insekte kako bi raširio svoje spore. Za dodatne informacije pogledajte Jetrenjače: Životni ciklus,
-
 Dijagram smjene generacija kod jetrenjača
Dijagram smjene generacija kod jetrenjača -
 Dijagram životnog ciklusa mahovine
Dijagram životnog ciklusa mahovine -
 Dijagram životnog ciklusa paprati
Dijagram životnog ciklusa paprati



U paprati i njihovim srodnicima, uključujući preslicu i konjski rep, uočljivija biljka u polju je diploidni sporofit. Haploidne spore razvijaju se u sorusima na naličju i rasipaju se vjetrom (ili u nekim slučajevima, plutajući po vodi). Ako su uslovi dobri, spora će klijati i izrasti u prilično neupadljivo biljno tijelo zvano protalus. Haploidni protalus ne liči na sporofita, pa kao takve paprati i njihovi srodnici imaju heteromorfnu smjenu generacija. Protalus je kratkotrajan, ali ima spolno razmnožavanje, proizvodeći diploidni zigot koji iz protalusa raste kao sporofit. Za dodatne informacije pogledajte Paprat#Životni ciklus.
-
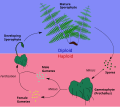
Dijagram smjene generacija kod paprati. -

Gametofit (protalus) Dicksonia sp. -

Sporofit Dicksonia antarctica. -

Naličje Dicksonia antarctica sa sorusima – strukturama za stvaranje spora




U grupi Spermatophyte, u sjemenjačama, sporofit je dominantna višećelijska faza; gametofiti su jako smanjeni i vrlo se razlikuju u morfologiji. Čitava generacija gametofita, s izuzetkom polenova zrna (mikrogametofiti), nalazi se unutar sporofita. Životni ciklus biljke cvjetnice (angiosperme), vrbe, detaljno je opisan u prethodnom odjeljku. Životni ciklus gimnospermi je sličan. Međutim, cvjetnice imaju i fenomen koji se naziva „dvostruka oplodnja“. Dva jedra sperme iz polenova zrna (mikrogametofit), a ne jedno, ulaze u arhegonij megagametofita; jedna se stapa s jedrom jajašca za formiranje zigota, a druga se s dva druga jedra gametofita, kako bi se formirao endosperm, koji hrani embrion koji se razvija.
Za dodatne informacije pogledajte Dvostruka oplodnja.
Evolucija dominantnosti diploidne faze
[uredi | uredi izvor]Predloženo je da je osnova za nastanak diploidne faze životnog ciklusa (sporofita) kao dominantne faze (npr. Kod vaskularnih biljaka) to što diploidija omogućava maskiranje ekspresije štetnih mutacija putem genetičke komplementacije.[23][24]
Prema tome, ako je jedan od roditeljskih genoma u diploidnim ćelijama sadržavao mutaciju koja dovodi do oštećenja u jednom ili više genskih proizvoda, ti bi nedostaci mogli biti nadoknađeni drugim roditeljskim genomom (koji ipak može imati svoje vlastite mane u drugim genima). Kako je diploidna faza postajala prevladavajuća, učinak maskiranja vjerovatno je omogućio da se veličina genoma, a time i sadržaj informacija, poveća bez ograničenja na to da se mora poboljšati preciznost replikacije DNK. Mogućnost povećanja sadržaja informacija po niskoj cijeni bila je povoljna jer je omogućila šifriranje novih prilagodbi. Ovo gledište je osporavano, a dokazi pokazuju da selekcija nije efikasnija u haploidnoj nego u diploidnoj fazi životnog ciklusa mahovina i angiospermi.[25]
-
 Životni ciklus angiospermi
Životni ciklus angiospermi -
 Vrh tulipanovog prašnika sa polenom (mikrogametofiti)
Vrh tulipanovog prašnika sa polenom (mikrogametofiti) -
 Biljne ovule (megagametofiti): Ovule golosjemenjača (lijevo) i angiospermi
Biljne ovule (megagametofiti): Ovule golosjemenjača (lijevo) i angiospermi -
 Dvostruka oplodnja
Dvostruka oplodnja




Slični procesi kod ostalih organizama
[uredi | uredi izvor]Rhizaria
[uredi | uredi izvor]Neki organizmi, koji su sada klasificirani u kladus Rhizaria i biljke u smislu kakav se ovdje koristi ne pokazuju smjenu generacija. Većina Foraminifera prolazi kroz heteromorfnu smjenu generacija između haploidnih “gamont“ i diploidnih “agamont“oblika. Jednoćelijski haploidni organizam je obično mnogo veći od diploidnog organizma.
Gljive
[uredi | uredi izvor]Miceliji gljiva su tipski haploidi. Kad se susretnu dva uparujuća micelija različitih vrsta, stvaraju dvije multijedarne ćelije u obliku kuglice, koje se spajaju preko „mosta za parenje“. Jedra prelaze iz jednog micelija u drugi, formirajući heterokarion (što znači "različita jedra"). Ovaj proces se naziva plazmogamija. Stvarno stapanje u diploidns jedra naziva se kariogamija i ne može se dogoditi dok se ne formiraju sporangije. Karogamija proizvodi diploidni zigot, što je kratkotrajni sporofit koji ubrzo prolazi kroz mejozu da bi formirao haploidne spore. Kad spore klijaju, razvijaju se u nov micelij.
Sluzave plijesni
[uredi | uredi izvor]Životni ciklus sluzavih plijesni je vrlo sličan životinjskom. Haploidne spore klijaju u tvorevine gomile ćelija ili myxamoebae. One se spajaju u procesu koji se naziva "plazmogamija" i "kariogamija" da bi se formirao diploidni zigot. Žigot se razvija u plazmodij, a kad ovaj sazri proizvodi, od jednog do mnogo plodnih tijela koja sadrže haploidne spore, ovisno o vrsti.
Životinje
[uredi | uredi izvor]Smjena između višećelijske diploide i višećelijske haploidne generacije se nikad ne susreće kod životinja. Kod nekih životinja postoji smjena između faza partenogeneze i seksualne reprodukcije (heterogamija). Obje faze su diploidne. To se ponekad nazivalo „smjena generacija“, ali je sasvim drugačija. U nekim drugim životinjama, kao što su insekti reda Hymenoptera (gdje spada i pčela), mužjaci su haploidni, a ženke diploidne, ali to je uvijek slučaj, umjesto da postoji smjena između različitih generacija.
Također pogledajte
[uredi | uredi izvor]Reference
[uredi | uredi izvor]- ^ Kerp, H.; Trewin, N.H.; Hass, H. (2003), "New gametophytes from the Lower Devonian Rhynie Chert", Transactions of the Royal Society of Edinburgh: Earth Sciences, 94 (4): 411–428, doi:10.1017/S026359330000078X Nepoznati parametar
|lastauthoramp=zanemaren (prijedlog zamjene:|name-list-style=) (pomoć) - ^ a b Svedelius, Nils (1927), "Alternation of Generations in Relation to Reduction Division", Botanical Gazette, 83 (4): 362–384, doi:10.1086/333745, JSTOR 2470766
- ^ Taylor, Kerp i Hass 2005
- ^ ""Plant Science 4 U". Pristupljeno 5. 7. 2016.
- ^ a b Bateman i Dimichele 1994, str. 403
- ^ a b Stewart i Rothwell 1993
- ^ a b c d Haig, David (2008), "Homologous versus antithetic alternation of generations and the origin of sporophytes" (PDF), The Botanical Review, 74 (3): 395–418, doi:10.1007/s12229-008-9012-x, pristupljeno 17. 8. 2014
- ^ Hofmeister, W. (1851), Vergleichende Untersuchungen der Keimung, Entfaltung und Fruchtbildildiung höherer Kryptogamen (Moose, Farne, Equisetaceen, Rhizocarpeen und Lycopodiaceen) und der Samenbildung der Coniferen (jezik: German), Leipzig: F. Hofmeister, pristupljeno 17. 8. 2014CS1 održavanje: nepoznati jezik (link)
- ^ Currey, Frederick (1862), On the germination, development, and fructification of the higher Cryptogamia, and on the fructification of the Coniferæ, London: Robert Hardwicke, pristupljeno 17. 8. 2014
- ^ Feldmann, J.; Feldmann, G. (1942), "Recherches sur les Bonnemaisoniacées et leur alternance de generations" (PDF), Ann. Sci. Natl. Bot., Ser. 11 (jezik: French), 3: 75–175, arhivirano s originala (PDF), 19. 8. 2014, pristupljeno 24. 9. 2019 Nepoznati parametar
|lastauthoramp=zanemaren (prijedlog zamjene:|name-list-style=) (pomoć)CS1 održavanje: nepreporučeni parametar (link) CS1 održavanje: nepoznati jezik (link), p. 157 - ^ Feldmann, J. (1972), "Les problèmes actuels de l'alternance de génerations chez les Algues", Bulletin de la Société Botanique de France (jezik: French), 119: 7–38, doi:10.1080/00378941.1972.10839073 CS1 održavanje: nepreporučeni parametar (link) CS1 održavanje: nepoznati jezik (link)
- ^ Schopfer, P.; Mohr, H. (1995). "Physiology of Development". Plant physiology. Berlin: Springer. str. 288–291. ISBN 978-3-540-58016-4. Upotreblja se zastarjeli parametar
|chapterurl=(pomoć) - ^ Guiry i Guiry 2008
- ^ Bateman i Dimichele 1994, str. 347
- ^ a b Shyam 1980
- ^ Watson 1981, str. 2
- ^ Kirby 2001
- ^ Watson 1981, str. 33
- ^ Bell i Hemsley 2000, str. 104
- ^ Watson 1981, str. 425–6
- ^ Watson 1981, str. 287–8
- ^ "Willows", Encyclopædia Britannica, XIX (11th izd.), New York: Encyclopædia Britannica, 1911, pristupljeno 1. 1. 2011
- ^ Bernstein, H.; Byers, G.S.; Michod, R.E. (1981), "Evolution of sexual reproduction: Importance of DNA repair, complementation, and variation", The American Naturalist, 117 (4): 537–549, doi:10.1086/283734 Nepoznati parametar
|lastauthoramp=zanemaren (prijedlog zamjene:|name-list-style=) (pomoć) - ^ Michod, R.E.; Gayley, T.W. (1992), "Masking of mutations and the evolution of sex", The American Naturalist, 139 (4): 706–734, doi:10.1086/285354 Nepoznati parametar
|lastauthoramp=zanemaren (prijedlog zamjene:|name-list-style=) (pomoć) - ^ Szövényi, Péter; Ricca, Mariana; Hock, Zsófia; Shaw, Jonathan A.; Shimizu, Kentaro K.; Wagner, Andreas (2013), "Selection is no more efficient in haploid than in diploid life stages of an angiosperm and a moss", Molecular Biology and Evolution, 30 (8): 1929–39, doi:10.1093/molbev/mst095, PMID 23686659 Nepoznati parametar
|lastauthoramp=zanemaren (prijedlog zamjene:|name-list-style=) (pomoć)
Bibliografija
[uredi | uredi izvor]- Barnes, R.S.K.; Calow, P.; Olive, P.J.W.; Golding, D.W.; Spicer, J.I. (2001), The Invertebrates: a synthesis, Oxford; Malden, MA: Blackwell, ISBN 978-0-632-04761-1 Nepoznati parametar
|lastauthoramp=zanemaren (prijedlog zamjene:|name-list-style=) (pomoć) - Bateman, R.M.; Dimichele, W.A. (1994), "Heterospory – the most iterative key innovation in the evolutionary history of the plant kingdom" (PDF), Biological Reviews of the Cambridge Philosophical Society, 69 (3): 345–417, doi:10.1111/j.1469-185x.1994.tb01276.x, arhivirano s originala (PDF), 15. 4. 2012, pristupljeno 30. 12. 2010 Nepoznati parametar
|lastauthoramp=zanemaren (prijedlog zamjene:|name-list-style=) (pomoć) - Bell, P.R.; Hemsley, A.R. (2000), Green Plants: their Origin and Diversity (2nd izd.), Cambridge, etc.: Cambridge University Press, ISBN 978-0-521-64109-8 Nepoznati parametar
|lastauthoramp=zanemaren (prijedlog zamjene:|name-list-style=) (pomoć) - Foster, A.S.; Gifford, E.M. (1974), Comparative Morphology of Vascular Plants (2nd izd.), San Francisco: W.H. Freeman, ISBN 978-0-7167-0712-7 Nepoznati parametar
|lastauthoramp=zanemaren (prijedlog zamjene:|name-list-style=) (pomoć)</ref>[1] --> - Guiry, M.D.; Guiry, G.M. (2008), "Cladophora", AlgaeBase, World-wide electronic publication, National University of Ireland, Galway, pristupljeno 21. 7. 2011
- Kirby, A. (2001), Ulva, the sea lettuce, Monterey Bay Aquarium Research Institute, arhivirano s originala, 16. 5. 2011, pristupljeno 1. 1. 2011</ref>
- Scott, Thomas (1996), Concise Encyclopedia Biology, Berlin: Walter de Gruyter, ISBN 978-3-11-010661-9
- Shyam, R. (1980), "On the life-cycle, cytology and taxonomy of Cladophora callicoma from India", American Journal of Botany, 67 (5): 619–24, doi:10.2307/2442655, JSTOR 2442655
- Sporne, K.R. (1974a), The Morphology of Angiosperms, London: Hutchinson, ISBN 978-0-09-120611-6
- Sporne, K.R. (1974b), The Morphology of Gymnosperms (2nd izd.), London: Hutchinson, ISBN 978-0-09-077152-3
- Stewart, W.N.; Rothwell, G.W. (1993), Paleobotany and the Evolution of Plants (2nd izd.), Cambridge, UK: Cambridge University Press, ISBN 978-0-521-38294-6 Nepoznati parametar
|lastauthoramp=zanemaren (prijedlog zamjene:|name-list-style=) (pomoć) - Watson, E.V. (1981), British Mosses and Liverworts (3rd izd.), Cambridge, UK: Cambridge University Press, ISBN 978-0-521-28536-0
- Taylor, T.N.; Kerp, H.; Hass, H. (2005), "Life history biology of early land plants: Deciphering the gametophyte phase", Proceedings of the National Academy of Sciences of the United States of America, 102 (16): 5892–5897, doi:10.1073/pnas.0501985102, PMC 556298, PMID 15809414 Nepoznati parametar
|lastauthoramp=zanemaren (prijedlog zamjene:|name-list-style=) (pomoć)
Vanjski linkovi
[uredi | uredi izvor]Generationswechsel. Spektrum, Heidelberg 1999.
| Discipline | |
|---|---|
| |
| Rast i habitus (izgled) | |
| Razmnožavanje | |
| Taksonomija biljaka | |
| Primjena | |
| |